6. BÖLÜM
FOSİL KAYDI ▬ MEMELİLERİN KÖKENİ
Giriş
“Memeli benzeri” sürüngenler, yüksek derecede çeşitlilik içeren, memelilerde bulunan çok sayıda karakteristiğe sahip, genellikle dağınık bir sürüngen grubudur. Evrimciler, evrimin gerçek olduğunu ve memelilerin sürüngenlerden geldiğini varsayarak, oldukça mantıklı bir şekilde, bu memeli benzeri niteliklerinin varlığının, memelilerin, memeli benzeri sürüngenlerin içindeki canlıların bir veya daha çok grubundan meydana geldikleri kuramına destek sağladığını kabul ederler.
Elbette ki yaratılışçılar bu varsayımları kabul etmezler. Onlar, omurgalıların büyük bir çeşitlilik içerdiğine işaret ederler. Bazılarının ağırlığı otuz gramdan daha azken, bazılarınınki ise birkaç yüz tondur. Hareket biçimlerinde önemli farklılıklar bulunanlardan bazıları sadece kara ile kısıtlıdırlar. Bazıları yalnızca denizde yaşarken, bazıları usta uçuculardır. Evrim olsun ya da olmasın; eğer farklı sınıflardan olan omurgalılar pek çok karakteristiği ortak olarak paylaşmasalardı, işte bu gerçekten şaşırtıcı olurdu.
Probleme geniş bir açıdan baktığımızda, kanıtların tümünün yaratılış görüşünün lehinde olduğunu söylemek zorunda kalıyoruz. Çünkü, fosil kaydında, omurgalılar ile omurgasızlardan ata olduğu varsayılan herhangi bir canlı arasında bağlantı kurmamızı sağlayabilecek tek bir kanıt parçası bile yoktur. Bu geçiş için, 100 milyon yıllık bir dönem varsayılmasına rağmen, tek bir geçiş formu bile bulunamamıştır. Eğer omurgalılar (açıkça görüldüğü gibi) evrimleşmemişlerse, evrim kuramı ölmüştür ve omurgalı grupları ya da başka toplulukların evrimi konusunda fikirler yürütmek çok saçmadır. Eğer probleme daha dar bir açıdan bakarsak, yani ilgimizi sürüngenler, memeli benzeri sürüngenler ve memeliler üzerinde yoğunlaştırırsak, her iki bakış açısını da destekleyen kanıtlar bulabiliriz.
Kanıtlara Evrimsel Açıdan Bakış
İlk önce, memelilerin sürüngenlerden evrimleştiği tahminini savunan kanıtları inceleyelim. Bunu yaparken, eğer kanıtlar olması gerektiği gibi evrim modeli sanısı içinde değerlendirilecekse, jeolojik sıra ve zaman aralıklarına bir evrimci gözüyle bakacağız. “İlkel” memeli benzeri sürüngenler, fosil kaydında Geç Pennsilvaniyen Döneminde, “sürüngen benzeri” sürüngenlerle eşzamanlı olarak ortaya çıkmışlardır. Başlangıçtan itibaren bu canlılar, bugün memelilerle ilişkilendirilen belirli niteliklere sahiptiler; fakat, ikincil damak ve çift art kafa kondili gibi diğer memeli benzeri niteliklerden yoksunlardı. Daha sonra Permiyen ve sonra Trias’ta, bu ve diğer memeli niteliklerine, büyük ölçüde farklılık kazanmış dişlere, alt çenenin diş kemiklerinde bir büyümeye ve diğer alt çene kemiklerinde büyüklük olarak bir azalmaya sahip olan “gelişmiş” memeli benzeri sürüngenler ortaya çıkmışlardır. Ve sonuçta, evrimsel jeolojik zaman ölçeğine göre, yaklaşık olarak Trias–Jura sınırında ya da 180 milyon yıl önce bir canlı ortaya çıktı. Bu canlı, memeli benzeri niteliklerinin çoğuna, tümüyle işlevsel sürüngen tipi (dördül eklemli) bir çene eklemine ve bununla birlikte bir memeli çene eklemine (squamosal-dişle ilgili) sahip bir canlıydı. Yani, evrimcilerin, ilk memeli olarak nitelendirdikleri bir canlıya sahiptik artık. Memeli benzeri sürüngenler konusundaki en kapsamlı inceleme, Tom Kemp1 tarafından 1982 yılında basılmış olmasına rağmen, hâlâ kanıtlara evrimsel açıdan bakış konusunda kusursuz bir kaynaktır. Kanıt olarak değerlendirmemiz boyunca kullanacağımız ana kaynak da bu kitaptır.
Kanıtlara Yaratılışsal Açıdan Bakış
Evrimciler ne zaman evrimin kanıtlarından söz etmek isteseler daima sürüngen–memeli dönüşümüne Arkeopteriks (sürüngen–kuş arası bir ara seviye olarak ileri sürülen canlı) ve at serilerini işaret ederler. Daha önceden de belirttiğimiz gibi, Gould ve Eldredge Arkeopteriks’i bir ara seviye olarak kabul etmemektedirler2 ve Eldredge, atların evrim geçirdiğine inansa bile, at fosillerinin farklı türlerin arasında hiçbir geçiş formu bulunmadığını söylemektedir.3 Buna göre, gerçekten de yüz milyonlarca yıl boyunca milyonlarca tür kademeli olarak evrimleşmişlerse, evrime kanıt konusunda acınacak derecede az kanıt bulunduğu açıkça görülmektedir. Eğer bu gerçekten olmuşsa, bugün müzelerimizin çok sayıda tartışmasız geçiş formu ile dolu olması gerekirdi. Soruya gerek kalmaz, şüpheye ihtimal verilmez, tartışmaya fırsat verilmez, hatta Yaratılış Araştırma Enstitüsü’nün kurulmasının anlamı olmazdı. Bu şüphe götürmeyen geçiş formlarının var olmaları gerekirken evrim davası, sürüngenlerin memelilere dönüşümü gibi çok az sayıdaki şüpheli örnekle kalakalmaktadır.
Memeli benzeri sürüngenler ve memeliler ile ilgili bir evrim ya da bir soy oluş ağacını kabul ettirme girişimlerinde evrimciler, bu canlıları bir evrim senaryosuyla birbirine bağlama konusunda neredeyse tümüyle benzerliklere güvenmektedirler. Kuramsal evrim basamakları için gerekli olan geçiş formlarının yokluğu nedeniyle bunu yapmak zorundadırlar. Bunu daha önce, ileri sürülen geçişle ilgili kanıtların incelenmesinde belirtmiştik. Bir canlının, ikinci bir canlı sınıfı tarafından sahip olunan bazı niteliklere sahip olması, mutlaka bu iki sınıf arasında bir geçiş olduğunu mu gösterir? Bu soruya olumsuz bir yanıt vermek için çok sayıda örnekten söz edebiliriz. Seymouria, amfibyumlar ve sürüngenlerde bulunan bazı niteliklere sahip bir canlıydı. Bu nedenle bu canlının, amfibyumlar ve sürüngenler arası bir “mükemmel geçiş formu” olması gerekir. Ancak bu canlı böyle bir ara form olamaz. Çünkü, ilk olarak Orta Permiyen’in başlangıcında ortaya çıkan bu canlı, Pennsilvaniyen Dönemi başlarında ortaya çıkmış olan sürüngenlerin atası olabilmek için evrimsel zaman ölçeğine göre en azından 20 milyon yıl daha gençtir.
Bir diğer örnek, yaşayan ördek gagalı ornitorenktir. Bu canlı, bir memelidir ama bir gagası ve perdeli ayakları vardır ve yumurtlama özelliğine sahiptir; buna ek olarak, sürüngen nitelikleri olarak isimlendirilebilecek başka niteliklere de sahiptir. Memeli, sürüngen ve kuş niteliklerini barındıran bu canlı belki de “ilkel” bir memeli olarak isimlendirilebilir. Memelilerin atası olabilmek için çok geç zamanda ortaya çıkmış olması nedeniyle, memelilerin atası olabilmesi mümkün değildir! Gerçekte, bu benzersiz yapısal özellik birleşimi, bu canlının, belirli bir omurgalı sınıfından meydana geldiğini ya da herhangi iki sınıf arasında bir geçiş formu olabileceğini ileri sürmeyi imkansız hale getirmektedir. Pek çok benzer örnekten söz edilebilir. İki farklı türe ait hayvanların sahip oldukları niteliklerin tek bir canlıda var olması, bu canlının, bu iki türün arasında bir geçiş formu olduğunu ya da bu türlerin, birbirleriyle genetik anlamda akraba olduklarını göstermez.
Memeli benzeri sürüngenler, varsayımsal olarak, çok önceki sürüngen benzeri formlardan memeli benzeri formlara ve son olarak da memelilere geliştiği için, sürüngenden memeliye gelişimin düzenli olacağı düşünülebilir. Aslında, memeli benzeri sürüngenler, sürüngen ve memelilerde bulunan özelliklerin bir karışım modeline sahiptirler. Örneğin Colbert şunu söylemiştir:
Theriodont (“gelişmiş” memeli benzeri) sürüngenler arasındaki memeli atalarını kesin bir çizgiyle belirlemek kolay değildir. Bazı theriodont’lar, belirli karakterler bakımından memelilere doğru büyük ilerlemeler göstermişler fakat diğer yönlerden bir derece ilkel kalmışlardır. Tüm theriodont’lar arasında gelişmiş ve sade karakterlerin karışımı öylesine çeşitlidir ki, herhangi belirli bir grubu işaret etmek ve bu grubu, memeliler yönünde en olumlu anlamda gelişen bir grup olarak tanımlamak imkansızdır.4
Diğer bir deyişle, memeli benzeri sürüngenler olarak isimlendirilen canlılardan biri, ikincil bir damak ve ayrılmış dişler gibi memelilere ait olduğu söylenen belirli niteliklere sahip olmakla birlikte ilkel sürüngenlere ait olduğu tahmin edilen niteliklere de sahip olabilir. Colbert’e göre bu “gelişmiş” ve “ilkel” özellikler karışımı, theriodont’ lar (“gelişmiş” memeli benzeri sürüngenler) için öylesine yaygındır ki tek birini bile, memelilere giden yolda gerçek bir ata olarak seçmek imkansızdır.
Üstelik, memeli benzeri sürüngenler genellikle hem sürüngenlere hem de memelilere ait olduğu düşünülen niteliklerden oluşan karışımlar oluşturmalarının yanı sıra, bu sürüngenlerin birçoğunun sahip olduğu bazı diğer yapılar, memeli ya da sürüngenler gibi yaşayan hiçbir dört ayaklıda bulunmamaktadır. Kemp şöyle der:
Fosil yapılarının ayrıntılı anlamda canlı yapılara benzemesi olası değildir ve pek çok farklılık, önemli işlevsel farklılıkla ilişkilidir. Gerçekte, pek çok durumda memeli benzeri sürüngenlerin yapıları yaşayan dört ayaklıların yapılarlarına benzerlik göstermemektedir.5
Gayet açıktır ki, bu tür canlılar, memelilerin (ya da bu anlamda, başka herhangi bir yaşayan canlının) ataları olamayacak kadar çok ileri derecede özelleşmiş canlılardır.
Gerçekte, sürüngenlerden memelilere doğru gittiği varsayılan evrimsel çizgideki boşluklar öylesine sistematik boşluklardır ki bir canlının doğrudan diğer bir canlının atası olabileceğini düşünmek imkansızdır.
Tüm memeli benzeri sürüngenler, Synapsida altsınıfında yer almaktadırlar. Evrimciler, tümü sonuçta ortak bir atayı paylaşan bu canlıların doğal bir gruplama olduğunu, ortak atanın sürüngen tarihinin çok erken dönemlerinde var olduğunu varsaymaktadırlar. Bunlar gerçekte, birbirlerinden farklı olan canlılardır. Tüm üyelerde var olan tek ortak belirleyici özellik, sadece bu grupta bulunan, şakak bölgesindeki tek bir yanal açıklığın varlığıdır. Memeli benzeri sürüngenler, iki takıma ayrılmaktadır. Bunlar, memeli benzeri sürüngenlerin en erken üyelerini oluşturduğu düşünülen Pelycosauria ve evrimciler tarafından daha ileri türleri oluşturduğu düşünülen Therapsida dır. Pelycosauria’ lar, Avrupa’da çok az sayıda bulunmalarına rağmen, yalnızca Texas kırmızı yataklarında (Alt Permiyen) bolca bulunmaktadırlar. Therapsid’ler çoğunlukla Güney Amerika, Rusya ve Güney Afrika’daki Karoo Süpergrubu’nda bulunmaktadırlar. Romer bu canlıların bu iki takıma ayrılmalarını, mantıksız fakat işe yarar olarak ifade etmektedir.6 Bu iki takım içinde, familya, cins ve türler, sürüngenlerin ortaya çıkış sırasını simgeleyen bir sırada düzenlenmiştir.
Bu genel sıra, bu canlıların kökenleri konusunda gerçek bir zaman ardışıklığını mı simgeler yoksa bu ardışıklık evrim hakkındaki önyargılı fikirlere uygun olsun diye mi planlanmıştır? En azından, bu canlıların pek çoğunun evrim kuramının isteklerine göre sıralandığı konusunda birkaç temel şüphe görülmektedir. Ve sonra bu dizilim kuramına kanıt olarak ileri sürülmüştür!
Yukarıda sözü edilen şüpheyi belgelemek amacıyla, literatürden birkaç alıntı yapılabilir. Kemp şöyle der:
Ayrıca kayıtlar coğrafik anlamda yarım yamalak kayıtlardır; hiçbir yer, memeli benzeri sürüngenlerin tarihlerinin nispeten küçük bir kısmından daha fazlasını sağlamamaktadır ve pek çok durumda bir bölge, tek bir çağa ait fosiller içerir. Benzer şekilde, synapsidlerin dünya çapında bir sınıflandırma grubu bulunmamaktadır. Yine de bunların en azından bazılarının canlıyken yayıldıklarına ilişkin pek şüphe yoktur.7
Oldukça açık görülmektedir ki, hiçbir bölge memeli benzeri sürüngenlerin sözde tarihinin küçük kısımlarından daha fazlasını içermediği için, değişik kısımlar, hayali bir evrim sırasına ya da dolaylı evrim temellerine dayandırılan tahminlerce belirlenen bazı tasarılara göre yan yana getirilmelidir.
Özellikle açık olan şey Kemp’in şu sözleridir: “Jeologlar, synapsidleri, içinde bulundukları kıtasal kayaların nispi yaşlarının birer stratigrafik göstergesi olarak da kullanmaktadırlar... ”8 Diğer bir deyişle, fosiller, bu durumda memeli benzeri sürüngen fosilleri, kayaların yaşlarının belirlenmesinde kullanılmaktadırlar. Fakat, eğer biz memeli benzeri sürüngenleri kayaların yaşlarını belirlemede kulanıyorsak, o halde, bu canlıların nispi yaşlarını nereden biliyoruz? Elbette ki, evrimcilerin benimsedikleri evrimsel tasarımdaki sıradan!
Aynı derecede açık olan bir şey, Romer’in sözlerindedir:
Sualtı aşamasına ilişkin bağlantı pek çok durumda olanaksız olsa bile, therapsid’lerin ve kapsanan diğer formların genel evrim hikayesi, taşınan pelycosaur yataklarının Erken Permiyen, Beaufort’taki Tapinocephalus bölgesi ve erken Rusya tortularının Orta Permiyen ve yine Beaufort’taki Endothiodon ile Cistecephalus bölgelerinin ve eşdeğerlerinin Geç Permiyen olarak düşünülmesini önermektedir. Olson, orta teriminin çıkarılmasını ve Rusya ile Afrika yataklarının tümünün (ve Amerika’da bulduğu Çift Dağı ya da Pease Irmağı bulguları) Geç Permiyen olarak isimlendirilmesini önermiştir. Bu bana son derece pürüzlü bir bölünme gibi geliyor ve alışıldık erken–orta–geç terminolojileri, geniş açılı evrimsel Permiyen resmiyle daha çok uyum içinde bulunuyor.9
Öyle görünüyor ki, Romer’in, bu çeşitli yatakları varsayımsal bir zaman dizisiyle düzenlemesi, “therapsid’lerin genel evrimsel hikayesi” ve evrimsel Permiyen resmine geniş bir bakış açısı” ile belirlenmiştir. Öyleyse, hiçbir şüphe yoktur ki, literatürde ve özellikle de ders kitaplarında sunulan memeli benzeri sürüngenlerin zaman ardışıklığı konusu, genellikle evrimsel beklentilerle bir uyum içinde bulunmaktadır. Yani bu sunumlar, yalnızca istenilen uyumun sağlanabilmesi için oluşturulmuşlardır.
Ancak, fosillerin yaşlarının belirlenmesinde, herhangi bir stratigrafik ilişkiden ya da evrimsel hikayeden tümüyle bağımsız bir yöntem olan radyometrik yaş tayini yönteminin kullanıldığı sık sık dile getirilmektedir. Wales, Swansea’daki University Yüksek Okulu’nda Jeoloji Profesörü olan Derek Ager bu tür iddialara öfkeli şekilde şu yanıtı vermiştir:
Bir jeolog olarak benim öfkelerim, David Challinor’un doğa tarihi müzeleri ile ilgili makalesiyle (New Scientist, 29 Eylül, 1983, s. 959) ve özellikle, yazdığı şu sözlerle doruk noktasına ulaşmıştır: “Başlangıçta, paleontologlar fosillerin yaşlarını, içlerinde bulundukları jeolojik katmanın kimliğinin saptanması yoluyla belirliyorlardı. Bugün bir fosilin yaşı, radyoaktif karbonun ya da radyoaktif potasyumun argona bozunmasının ölçülmesi yöntemiyle belirlenmektedir”... 19. yy’ın başlangıcından, William Smith’ten bu yana fosiller, içlerinde bulundukları kayaların ilişkilendirilmesi ve yaşlarının belirlenmesinde en iyi ve en doğru gösterge olmuştur... İzotopik bozunmanın ölçümü ve fizikçilerin kazandığı övgüye gelince, kan kaynamaya başlar. Tabi ki bu tür çalışmalar milyonlarca yıla dayanan, büyük hata paylarıyla dolu tarihler veririler... Fosillerin yaşlarının belirlenmesinde kullanılmakta olan hiçbir radyoaktif bozunma örneği düşünemiyorum.10
Görülüyor ki, kayaların yaşlarının belirlenmesinde, radyometrik yaş tayini yöntemleri değil, fosiller kullanılmaktadır. Bu durum bizi, şu sorunun sorulduğu bir çemberin içine geri götürecektir: Fosillerin yaşlarını nasıl belirliyoruz? Aslında en sonunda, bütün yaşlar varsayımsal evrimsel sıraya göre belirlenmektedir.
Bu bölümün geri kalan kısmı, sürüngenler ve memeliler arasındaki büyük farkların değerlendirilmesi ve devamında, “Veriler, kökenlerle ilgili hangi modele, evrime mi yaratılışa mı, daha iyi uymaktadır?” sorusuna cevap bulunabilmesi amacıyla memeliler ve memeli benzeri sürüngenlerin fosil kaydının dikkatli bir incelenmesine adanmıştır.
Sürüngenler ve Memeliler Arasındaki Fiziksel ve Fizyolojik Farklılıklar
Bir sürüngenin, bir memeliye dönüşümünü gerektiren fizyolojik ve beraberindeki morfolojik değişimler çok derindir. İngiliz yaratılışçı bilim adamı Douglas Dewer’in kendisinin bile tam olmadığını söylediği listede, memeliler ile sürüngenler arasında 21 fark vardı.11 Kemp, memelilerle sürüngenler arası kuramsal ara seviyelerin bir listesini ve geçiş formlarının yokluğu nedeniyle evrimsel değişimlerin kademeli mi yoksa belirli miktarlarda mı gerçekleştiğini bilmenin zorluklarını sunmakta ve bu konularda tartışmaktadır. Ondan sonra şöyle demektedir:
Sonuçta, elimizde bulunan soy oluşsal çözünürlükle, her sistemin evriminin kademeli olduğu söylenebilir. Değişik sistemlerin evrimleri arasında görünürde şöyle bir ilişki vardır; değişik sistemlerdeki değişimlerin, diğer sistemlerdeki değişimle birlikte olma eğilimi vardır... Fosil kaydında hiç gözlenmeyen ya da olsa bile sadece ilişkileri kesin olmayan iskeletsel niteliklerde gözlemlenebilen başka memeli nitelikleri vardır. Kalbin yapısı, iki misli kan dolaşımı, böbrek ve onun özelleşmiş fizyolojisi, saç, emzirme ve sıcaklık fizyolojisi gibi özellikler, memeli kökeninin doğasını anlamada birinci derecede önem taşımaktadırlar. Kemik özelliklerine bakılarak, büyük olasılıkla bu yumuşak yapıların erken memeliler ve memeli benzeri sürüngenler yoluyla dereceli biçimde evrimleştiği düşünülmektedir. Fakat, böylesi bir hipotezi detaylı inceleyebilmek için, tüm bu çeşitli niteliklerin görevlerinin, birbirleriyle ve çevreleriyle nasıl bağlantılı oldukları hesaba katılmalıdır.12
Kemp, karada yaşayan bir canlının karşı karşıya kaldığı başlıca üç problem sıralamış ve memeliliğin kökeni konusundaki bu problemlerin her birini tartışmıştır. Bu üç ana problem, sıcaklık kontrolü, kimyasal kontrol ve hareket kontrolüdür. Memeli fizyolojisi, sabit ve nispeten yüksek bir vücut sıcaklığını korumaya elverişli olmalıdır. Endotermi olarak isimlendirilen bu özellik, soğuk kanlı (ektotermik) bir sürüngendekinin yaklaşık yedi katı olan bir hücresel metabolizma oranında ısı üretimi yoluyla gerçekleşmektedir. Endotermi, memelilerde bulunan fakat sürüngenlerde bulunmayan mükemmel biçimde ayarlanmış oldukça karmaşık bir biyolojik organizasyon gerektirir. Bir sürüngenin bir memeliye dönüşümü esnasında nelerin evrim geçirmesi gerektiği konusunda Kemp şöyle der:
... ne hareket düzeylerinin farklılaşması sırasında meydana gelen metabolik ısı üretimi oranındaki değişikliklerin ne de çevre sıcaklığı değişimlerinin vücut sıcaklığının değişimine neden olmaması için, mükemmel bir sıcaklık düzenleme kontrol mekanizmasının varlığı gereklidir. Bu yüzden saçlar, ter bezleri ve deride bulunan özelleşmiş kan damarları evrimleşmelidir. Memeli biyolojisinde endoterminin işleviyle ilgili daha dolaylı fakat aynı derecede önemli başka birkaç durum daha vardır. Hareket üyeleri, on kat artan yiyecek ihtiyacını karşılamak için canlıyı taşıyabilecek nitelikte yapılara dönüşmelidirler. Beslenme yapıları, yine bu büyük oranda besin alımını gerçekleştirmeli ve eğer, yiyeceklerin parçalanması, sadece bağırsaklarda gerçekleşen süreçlere bırakılırsa, çok çok uzun bir süreç olan yiyeceklerin sindirilmesi işlemine de yardımcı olmalıdır. Dış ortamla vücut arasında daha büyük oranda gaz girişçıkışı olacağı için diyafram gereklidir. Yüksek sıcaklıktan ve nefes alma oranının artmasından kaynaklanan su kaybının artma tehlikesi, böbreklerin mücadele etmesini gerektirir. Son olarak, bu faaliyetleri kontrol ve organize etmek üzere duyu organları ve merkezi sinir sisteminin ayarlanması gereklidir.13
Memeli vücudunun izolasyonu, kılları gerektirir. Sürüngen pulları ile tüyleri arasındaki büyük farklılık ve onların gelişim biçimleri daha önce tartışılmıştır. Kıllar, tüylerde olduğu gibi, foliküllerden oluşmaktadır ve böylece pullardan tamamıyla farklı bir gelişim biçimi göstermektedir. Evrimciler, sürüngen pullarının memeli kıllarına dönüşümü probleminin, sürüngenlerde gerçekleşen rastgele ve tesadüfî genetik hatalar sonucu “çözümlendiğine” bir şekilde inanmak zorundadırlar.
Kemp, “memeliler tarafından çözümlenmiş ikinci büyük karasal problem” şeklinde ifade ettiği kimyasal kontrolün evrimleşmesini tartışmaktadır. Evrimciler evrimsel süreçlerde “çözümlendi” gibi kelimeler kullanarak sanki, bilinçsiz evrim sürecinde, zekâ, plan ve deney varmış gibi konuşmaktadırlar. Tüm dört ayaklılarda olduğu gibi memelilerde de aşılması gereken problem, su kaybına eğilimli olma durumudur. Bu problemin aşılmasıyla ilgili ana organ, böbrektir. Kemp bu organı, memelilerde, diğer omurgalılardakinden çok daha özenli bir çalışma sistemine sahip bir organ olarak nitelendirmiştir. Memeli böbreklerinin karmaşıklığını açıklarken Kemp şöyle der:
Böbreklerde bulunan kan damarlarında, kan basıncı yüksektir ve böbrek tüplerinin sayısı fazladır. Bu yüzden memeli böbrekleriyle ilgili ilk önemli nokta, kanda çok yüksek bir süzme oranının olmasıdır. İkinci önemli nokta, ana su koruma yöntemi olan yoğun ve hipertonik idrar üretimiyle ilgili Henle Kulbu’nun çok uzun oluşudur. Üçüncü önemli nokta, hipertonik idrarı üreterek, canlının bunu sıvı halde vücuttan atabilmesi için, vücutta yeterli miktarda suyun korunmuş olmasıdır. Bu yüzden suyun vücuttan atılması, plazma içindeki iyonların ve diğer çözülebilir maddelerin miktarlarının çok iyi düzenlenmesine fırsat verir. Uygun oranlarda salgı salgılanması ve böbrek tüplerinden akan sıvı maddenin geri emilimi yoluyla, kandaki iyon ve molekül düzeyleri sabit halde korunabilmektedir.14
Kemp, ince ayarlanmış bir hemostatik sistemle bütünleşmesi gereken enzim kontrollü tepkimelerin çokluğundan bahseder. Sonra sözlerine şöyle devam eder:
Kalp ve dolaşım sistemi, böbrekler tarafından ihtiyaç duyulan yüksek kan basıncını üretebilecek şekilde tasarlanmalıdır. Ayrıca, kontrol edilen maddelerin her birinin miktarını saptamak ve de böbrek tüplerindeki geri emilimi ve uygun oranda salgılamayı başlatmak için, karmaşık bir endokrin sistemin varlığına ihtiyaç vardır.15
Memeliler tarafından yapılan hareket kontrolü, yani çok çeşitli kara yüzeylerinde gezebilme konusunda Kemp şöyle demektedir:
Memeli bacaklarının hareket etme açıları geniştir; bu nedenle de düzensiz zeminlerin üstünden geçebilir ya da engelleri atlatabilirler. Bacaklar uzun, incedirler; hızlı hareket yeteneği kazandırırlar ve ayakların, zeminde hayvanın simetri eksenine çok yakın bir yerde bulunması, hayvanların manevra yapabilmelerini ve daha çevik olabilmelerini sağlar. Hareket sistemindeki bu geometrik özelliklerin yanı sıra memeliler, onlara devamlı ve hızlı hareket edebilme yeteneği kazandıran kaslara sahiptirler. Böylece memelilerin hareket açısından sürüngenlere göre önemli derecede daha hızlı ve etkin olmalarının yanı sıra, memeliler hızlı hareketlerini kendileriyle aynı büyüklükteki bir sürüngenden çok daha uzun bir zaman süresi boyunca da devam ettirebilirler.
... Bacaklarda gerçekleşen değişimlerin yanı sıra, hareket sisteminde de şunlar gereklidir: artan besin ve oksijen ihtiyacının karşılanması, kontrol sağlayan karmaşık bir duyumsal ve merkezi sinir sisteminin gelişmesi ve belki de uzun süren hareketler için daha yüksek bir metabolizma hızına sahip olunması.16
Kemp daha sonra, memelilerde var olan, dış ortamdaki iniş–çıkışlara karşı koyabilmek için, hayvanın kendi iç ortamında düzenlemeler gerçekleştirebilme kabiliyeti olan homeostaz (homeostasis) kavramını tartışmaktadır. Kemp, memelilerde homeostazın korunması ile ilgili olan süreçleri ve yirmi yedi yapının bütününü resimlemiştir. Kemp, homeostazın hiçbir bölümünün bağımsız biçimde işlev göremeyeceğini ve var olan şeyin birleşmiş tek bir homeostatik mekanizma olduğunu vurgulamıştır.
Son olarak Kemp şöyle der:
Sonuç olarak, memeli biyolojisinin esası, çeşitli yapılar ve işlevsel süreçlerin çok yüksek karmaşıklık derecesi ve içsel bütünlüğüdür.17
Daha önce Kemp, memelilerin nitelikleri, özellikle homeostazın devam ettirilmesiyle ilgili yapı ve süreçleri, oldukça karmaşık ve tümüyle bütünleşmiş olduğu için, bu niteliklerin evrimsel kökenlerinin kademeli değişimler geçireceğini vurgulamıştı. Bunlardan hiçbiri bir diğerinden bağımsız olarak ortaya çıkamazdı.18 Kemp bunu, daha sonraki bir bölümde şu sözlerle yeniden vurgulamaktadır:
Fosil kaydının, homeostazın memeli seviyelerine doğru ilerleyen evriminin, organizmaların tüm kısımlarında aynı anda gerçekleştiği görüşünü desteklediği belirtilmiştir. Hiçbir yapı ya da işlev, diğer tüm özelliklerde gerçekleşen uygun değişimlere ayak uydurmadığı sürece çok büyük ölçüde evrimleşememektedir.19
Kemp, bir sürüngenin bir memeliye dönüşümü esnasında homeostazın sürdürülebilmesi için gerekli olan içsel değişikliklerin kademeli ve diğerlerine uyum sağlar biçimde olması gerektiği gibi, morfolojik değişimlerin de uyumlu ve kademeli olması gerektiğini söyleyerek açıklamalarına devam etmektedir. Kemp şöyle demektedir:
Disinodontları (Dicynodont) örnek olarak alırsak, bu canlıların otçul özelleşmeleri, dişler ile sert diş tabakalarının yer değiştirmesini, çene kaslarının yönlenmesini, çenenin eklem yapısındaki değişimleri ve kafatası ile alt çenenin biçimlerinin büyük oranda yenilenmesini gerektirmektedir. Ayrıca, uygun gezme kabiliyeti ve merkezi sinir sistemine bağlı düzenlemeler ile davranışlar gerekmektedir. Bu özelliklerin hiç birisi, diğerleri ile birlikte bulunmadıkça hiçbir değer taşımamaktadır. Bu yüzden, disinodont organizma türünün evrimleşmesi için, bunu, gelişen her özelliğin kademeli olduğu ve diğer özelliklerde gerçekleşen değişimlerin eşliğinde geliştiği, birbiriyle bağlantılı bir gelişim süreci izlemelidir.20
Memeli Benzeri Sürüngenlerin Fosil Kaydı
Kemp kendi mantığına göre haklı olarak, bir sürüngenin bir memeliye dönüşebilmesi için, fizyolojide çok sayıda içsel değişimin ve morfolojide çok sayıda dışsal değişimin kademeli olarak gerçekleşmesi gerektiğini iddia etmektedir. Eğer evrim gerçekse, fosil kaydı tüm türler, cinsler, familyalar ve takımlar için kademeli bir köken belgelemelidir.
Birçok durumda bizler, çok farklı türlerdeki memeli benzeri sürüngenlerin evrimleşmesi esnasında bıraktıkları geçiş formları aracılığıyla her farklı türün kökenini izleyebilmeliyiz. Böylece sadece memeli benzeri bir canlının değil, %100 memeli olan bir canlının var olduğu son aşamaya ulaşabilmeliyiz. Bundan sonra, fosilleşmiş geçiş formları aracılığıyla, bu ilkel memeliden ortaya çıkan otuz iki memeli takımının –kemirgenler, tek sayıda parmaklı toynaklılar (perissodactyl), çift sayıda parmaklı toynaklılar (artiodactyl), balinalar, yarasalar, maymunlar ...vb.– her birinin kademeli kökenini izleyebilmeliyiz.
Fosil kaydı, ne evrim temellerinin dayandırıldığı geçiş formlarına ne de kademeli değişime kanıt sağlamaktadır. Kemp, kitabının en başında, sürüngenlerden memelilere dönüşümün, bir omurgalı sınıfından diğer bir omurgalı sınıfına dönüşümün nasıl olduğunu gösteren fosil kaydı tarafından çok iyi belgelenmiş tek örnek olduğunu iddia ettikten hemen sonra şunu itiraf etmektedir:
Elbette ki synapsid fosil kaydında pek çok boşluk vardır; bilinen farklı gruplar arasındaki ara seviye formları, neredeyse her zaman kayıptırlar. Ancak, bilinen gruplar yeterince ortak özelliğe sahip olduğu için kuramsal ara seviyeleri tasarlamak mümkündür.21
Kemp, sürüngenden memeliye dönüşümün, evrimde en iyi biçimde belgelenmiş durum olduğunu iddia edip, geçiş formları neredeyse daima kayıp oldukları için, kuramsal geçiş formlarının oluşturulması gerektiğini itiraf etmek zorunda kalmıştır! İlk memeli benzeri sürüngenler Üst Pennsilvaniyen (Karbonifer) kayaları arasında, sözde yaklaşık 350 milyon yıl önce ortaya çıkmışlardır ve Trias sonunda soyları tükenmiştir. Buna göre evrimciler, memeli benzeri sürüngenlerin, memeli konumuna erişmeden önce, neredeyse 200 milyon yılını evrimleşmekle geçirdiklerine inanmaktadırlar. Bu uzun zaman sürecinde milyarlarca geçiş formu yaşamış ve ölmüş olmalıdır. Müzelerimizdeki raflarda, binlerce gerçek geçiş formu bulunmalıdır. Eğer bu canlılar gerçekten evrimleşmiş olsaydı, kuramsal ara formlarda çare aramak kesinlikle gerekli olmazdı.
Geçiş formlarının yokluğu, türler, cinsler, familyalar ve takımlar gibi tüm sınıflandırma birimlerinde var olan bir gerçektir. Kemp şöyle demektedir:
Tür ve cins gibi alt sınıflandırma birimlerinde yer alan boşluklar, memeli benzeri sürüngenlerin fosil kaydının hemen hemen tümünde vardır. Uygun şekilde belgelenmiş durumların hiç birinde, türden türe geçiş aracılığıyla bir cinsten başka bir cinse geçişi izlemek mümkün değildir.22
Kemp, türler ve cinsler arası geçiş formlarının yokluğunu, küçük popülasyonlarda tür seviyesinde gerçekleşen evrimin hızlı bir şekilde gerçekleştiği fikrine dayandırmaktadır. Kemp, Niles Eldredge ve Stephen Jay Gould tarafından ortaya atılan sıçramalı denge evrim kuramını kabul etmektedir. Bu konu, daha sonraki bir bölümde tartışılacaktır. Sıçramalı denge evrim kuramı, türler arası geçiş formlarının yokluğu ile ilgili olarak ortaya atılmıştır fakat bu kuramın, cinsler, familyalar, takımlar, sınıflar ve filumlar hakkında söyleyebilecek hiçbir şeyi yoktur. Temelde farklı türler içeren familyalar, takımlar ve daha yüksek taksonlar arasındaki geçiş formlarının yokluğunu açıklamak için, kanıtlanmamış (ve kanıtlanamayan) bu hipoteze atıfta bulunmak tamamıyla geçersizdir.
Kemp, geçiş formlarının yokluğunun tüm seviyelerde açık olduğunu kabul etmekte ve şöyle demektedir:
Daha yüksek taksonlar, familyalar ve hatta takımların, bir kitle yok oluşundan hemen sonra, tüm özellikleri az ya da çok gelişmiş biçimde aniden ortaya çıkması, çok hızlı gerçekleşmiş bir evrimi işaret eder. Bundan sonra nesilde, çoğu zaman cins seviyesini aşmayan ama en fazla alt–familya seviyesinde seyreden, daima daha düşük oranlı morfolojik değişimler meydana gelir. Bu gözlemin bir ayak izi olması ve aslında yeni taksonların fosil kaydında ortaya çıkmadan önce uzun bir tarih içerisinde kademeli olarak öznitelik kazanmış olmaları olasıdır. Ancak, hiçbir durumda tek bir örnekte bile böylesi uzun bir tarih bilinmemektedir; o yüzden yüksek oranlardaki morfolojik evrimin, özellikle bir kitle yok oluşunun ardından gerçekleştiğini kabul etmek çok daha mantıklıdır.23
Kemp’in, tür ve cinslerde olduğu gibi, familya ve takım seviyelerindeki memeli benzeri sürüngen temsilcilerinin de tam halini almış ve özellikleri hemen hemen tamamıyla gelişmiş bir biçimde ortaya çıktıklarını kabul etmek zorunda kaldığına dikkat ediniz. Kemp, bunun, yüksek oranlardaki morfolojik evrimin özellikle bir kitle yok oluşundan sonra gerçekleştiğini kabul etmenin mantıklı olduğu hızlı bir evrimi işaret ettiğini söylemektedir.
Kemp’in inandığı şey, fosil kaydının, sürüngen–memeli dönüşümünün değişik aşamalarını simgeleyen memeli benzeri sürüngenlerin, her aşaması memeli benzeri niteliklere doğru artış gösteren ani bir ortaya çıkışla var olduklarını gösterdiğidir. Bir aşamanın bir diğerine kademeli biçimde dönüşümünü belgeleyen hiçbir geçiş formu olmaksızın, bu aşamaların fosil kalıntıları tam halini almış bir biçimde görünmektedir. Bir aşama içinde, sonraki aşamaya gelinceye kadar da pek bir değişim görünmemektedir.
Darvinci olmayan bu evrim fikrini açıklayabilmek için Kemp, şu önermede bulunur: Memeli benzeri sürüngenlerin gelişimlerinin her aşamasında hızlı bir yayılma vardı; ve bunu takiben kitlesel yok oluşlar ortaya çıktı. Her yok oluşu, hayatta kalan tek bir nesille başlayan bir yayılım izledi. Bu yayılım, sonuçta yine de kitlesel yok oluşu yaşayacak olan bir sonraki aşamaya kadar devam etti. Her yayılımın daha gelişmiş memeli benzeri canlılarla sonuçlandığı yayılım ve kitle yok oluşları dizisi, sonuçta, memelilerinin kökeninin ortaya çıkışıyla doruk noktasına ulaştı. Kemp şöyle der:
Yeni nesiller fosil kaydında ilk ortaya çıktıklarında hemen hemen tamamıyla gelişmiş halde ve daha sonra nispeten değişmeden kalmışlardır.24
Kemp bize, bu evrimsel gelişimin üç evrede gerçekleştiğini söyler. İlk evre, Üst Pennsilvaniyen ve Alt Permiyen’de pelycosaur’ların ortaya çıkışlarıyla sonuçlanmıştır. Kemp şöyle der:
Tüm pelycosaur’lar arasındaki yapısal benzerlikler öylesine çoktur ki, bu canlıların, grubun başlıca özelliklerini çoktan evrimleştirmiş olan ortak bir atayı paylaştıklarına hiç şüphe yoktur.25
Pelycosaur’ların ortak bir atadan evrimleştikleri iddiasının, sadece, belirli benzerliklere sahip olmanın ortak bir atayı gerektirdiği varsayımına dayandırıldığına dikkat ediniz. Gerçek kanıtlar bu fikre zıttır; çünkü, varsayılan ortak ata ilk ortaya çıkışında bu başlıca grup özelliklerinin tümüne zaten sahiptir. Ama fosil kaydında, kuramsal ortak atanın evrimsel kökenini ya da kuramsal ortak atanın farklı tür pelycosaur’lara evrimleştiğini belgeleyen hiçbir geçiş formu, tek bir tane bile, bulunamamıştır.
Şüpheli bir akrabalık taksonu olan Cotylosauria, sözde ilkel sürüngen olan dört ayaklıların birçok çeşidi için sıkça kullanılmaktadır. Böylece tanım olarak Cotylosaur’lar, ata olan sürüngenlerdir. Durum böyle olduğu için evrimciler, pelycosaur’ların bir cotylosaur’dan evrimleştiğini varsaymak zorundadırlar. Bunun için Romer ve Price şöyle demektedirler: “Pelycosaur’ların, Cotylosaur’ların soyundan geldiği kesin görünmektedir.”26 Ancak biraz sonra şöyle demektedirler: “pelycosaur atalarını cotylosaur takımı içinde aramaya yönelik bir girişim daha zordur.” Eleme süreci sonunda, cotylosaur’lar arasında, captorhinomorph’ların, pelycosaur’ların atalarını içermesi gerektiği kararına varırlar fakat şunu da itiraf ederler: “Pelycosaurları diğerlerinden ayıran özellikleri bulmaya yönelik girişimler, düş kırıklığına uğratıcı girişimlerdir.”
Böylece pelycosaurlara özgü nitelikler taşıyan sürüngenler, fosil kaydında birdenbire ve tam halini almış bir biçimde ortaya çıkmışlardır. Cotylosaurlar arasında bu benzersiz özelliklerin herhangi birine sahip herhangi bir olası ata bulunamamıştır; bu yüzden evrimciler, sadece pelycosaurların, cotylosaurlardan evrimleştiğini varsaymışlardır; çünkü önerebilecekleri daha iyi bir ata yoktur.
Pelycosauria hakkındaki kapsamlı inceleme yazılarında Romer ve Price, memelilerin evrimsel kökeninde bir sonraki aşamayı oluşturan Therapsida’nın atalarının, Pelycosauria’nın içinde bulunduğu konusunu ısrarla tartışmışlardır.27 Romer ve Price, therapsid’ler ile pelycosaur’lar arasında olduğuna inandıkları sayısız benzerliği anlatmış ve dahası, Sphenacodontidae’de yer alan pelycosaur’ların therapsid’lerin atası olduklarını ileri sürmüşlerdir. Sphenacodontid’ler ve therapsid’lerin iskeletlerinin uzantıları arasındaki benzerliklerle ilgili olarak Romer ve Price’in en fazla söyleyebileceği, sphenacodontid’lerin iskelet uzantılarının “en az birkaç ayrıntıda therapsid özelliklerinin başlangıcını işaret ettiği”dir.28 Eksen iskeleti hakkında şöyle derler: “Eksen iskeleti, gruplar arası yakın bir genetik bağlantı kurmak için güçlü bir sav oluşturmamakta fakat diğer yandan hiçbir engel de getirmemektedir.” Yani, therapsid’lerle sphenacodontid’lerin eksen iskeletlerinde ve iskelet uzantılarında onları birbirine bağlayabilecek hiçbir gerçek benzerlik bulunmamaktadır. Ancak Romer ve Price şuna inanmaktadırlar: “Kafatası yapılarındaki benzerlikler çok sayıda ve dikkate değerdir.”29 Fakat bu benzerliklerin bile, alışkanlıklarla ya da paralelliklerle ilgili olabileceğini söylemektedirler. Daha iyi ata adaylarından yoksun olan Romer ve Price şu sonuca varırlar: “Bütün ihtimaller göz önüne alındığında, memeli benzeri sürüngenler, sphenacodontid pelycosaur’larının soyundan gelmişlerdir.” Ancak, bunun hemen arkasından şöyle derler:
Pelycosaur’lar, pek çok iskelet özelliği açısından oldukça ilkeldirler; bundan dolayı, pek çok bakımdan hemen hemen sonraki herhangi bir sürüngenin morfolojik öncüsü olabilirler.30
Pelycosauria sphenacodont’gillerinin, therapsid’lerin atası olduğu fikri ayrıca Hopson31 ve Kemp tarafından da desteklenmektedir.
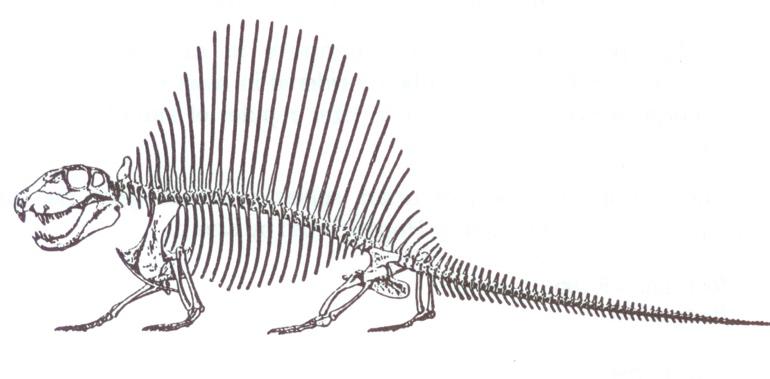
Dimetrodon, büyük bir ihtimalle ısı–değiştiricisi olarak görev yaptığına inanılan, yelken benzeri geniş yapılar oluşturan sırt ipine bağlı çok uzun dikenlere sahip çok sayıdaki sphenacodont’gillerden biriydi (Şekil 10). Bu kocaman dikenlerin, kademeli evrimsel kökenini gösteren hiçbir ara fosil bulunmamaktadır. Sphenacodon, sphenacodont’gillerin, en sade üyelerinden biriydi (Şekil 11). Therapsid’lerin kesin atasının hangi canlı olduğuna dair hiçbir öneri ortaya atılmadı.
Açıkça görülmektedir ki, pelycosaur’ları, therapsid’lerle ilişkilendirme tartışmaları son derece zayıftır ve sadece belirli benzerliklere dayanmaktadır. Pelycosaur’lar ve therapsid’ler arasında gerçek evrimsel zincir halkaları oluşturabilecek hiçbir geçiş formu bulunmamaktadır.
Kemp’e göre, pelycosaur’lardaki (bir veya birkaç nesil dışında) kitlesel yok oluşu, Geç Permiyen’de, cynodont dışındaki therapsid’lerin hızlı yayılımı izlemiştir ve bu yayılım tümüyle farklı memeli benzeri sürüngenlerden oluşan bir dizi şeklinde aniden ortaya çıkmıştır.32 Rusya, Güney Afrika ve Kuzey Amerika’da bulunan fosillerle ilgili olarak Kemp şöyle demektedir:
Bu üç kıtanın pelycosaur dışındaki memeli benzeri sürüngenlerin daha gelişmiş olanlarının tümü, Therapsida’nın üyeleridirler. İlk ortaya çıktıklarında bile, birçok farklı türe ayrılmışlardı; fakat, pelycosaur durumundan evrimleşen ortak özellikleri, Therapsida’ların tek bir kuramsal pelycosaur atanın soyundan geldiğini ve tek sınıflı bir grup oluşturduğunu göstermektedir.33
Şekil 10. Dimetrodonmilleri’niniskeletselgösterimi
(RomervePrice’den, s.337).
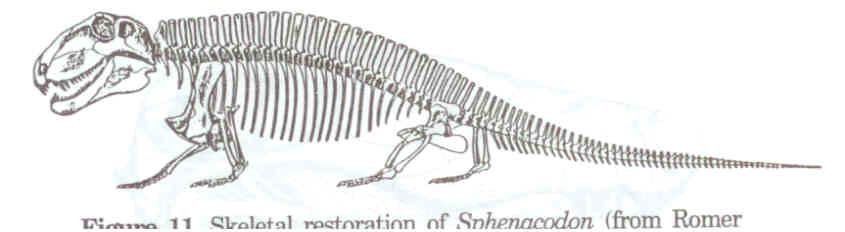
Şekil 11. Sphenacodon’un iskeletsel gösterimi
(Romer ve Price’den, s. 324).
Cynodont dışındaki therapsid’lerin tüm temel nitelikleri tamamlanmış biçimde aniden ortaya çıktıklarının vurgulanması gerekmektedir. İlk ortaya çıktıklarında, çok sayıda farklı tür oluşturmuşlardı ve bundan dolayı kuramsal bir pelycosaur’un ortak bir ata olarak önerilmesi gerekmektedir. Bu çok sayıdaki farklı türü birbirine ya da onları kuramsal bir pelycosaur ataya bağlayan hiçbir ara seviye, hiçbir geçiş formu bulunmamaktadır. Eğer bu canlılar ayrı ayrı yaratılmışlarsa, beklenen kanıt, kesinlikle bu kanıttır; fakat bu kanıt, evrim kuramının dayandırıldığı tahminlerle şüphesiz bir zıtlık içinde bulunmaktadır.
Cynodont dışındaki therapsid’ler, büyüklük, morfoloji ve beslenme alışkanlıkları bakımından çok farklıydılar. Bazıları etçil, bazıları otçuldu. Bazıları oldukça büyükken, bazıları çok küçüktü. Gorgonopsid’ler, “kılıç dişliler” olarak isimlendirildiler çünkü, hiçbir geçiş formu olmaksızın, tam anlamıyla bir gorgonopsid niteliği olan büyük köpek dişleriyle ortaya çıkmışlardır.34 Therocephalia grubundan Euchambersia, görünüşe göre, zehirli bir ısırığa neden olan yılan benzeri uzun sivri dişlere ve zehir bezelerine sahipti.35 Theriognathus gibi bazı whaitsiid therocephalia’ların hiçbir köpek dişi yoktu (Şekil 12).36
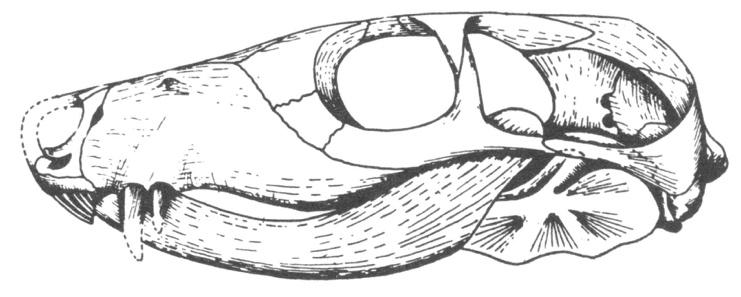
Şekil 12. Whaitsiid Therocephalia’larından Theriognathus.
(Brink, Paleontological africanus 4:97-115, 1956).
Therocephalia üyeleri, fosil kaydında ilk ortaya çıktıklarında tüm ayırt edici niteliklere sahiplerdi. Kemp onlarla ilgili olarak şöyle demektedir:
Şimdiye kadar tartışılmış olan hiçbir therapsid türünde paylaşılan ortak ata nitelikleri keşfedilmemiştir. Bu yüzden bunların diğer gruplarla olan akrabalıkları karanlıktadır.37
Therocephalia’lar, ilk temsilcilerinde tüm ayırt edici nitelikler tamamlanmış olarak ortaya çıktıkları ve bu ayırt edici niteliklerin hiçbiri memeli benzeri sürüngen therapsid’lerinin hiç birinde bulunmadığı için, evrim kuramının ihtiyaç duyduğu evrimsel zincir halkalarının hiç birinin ortada olmadığı ve bu canlıların diğer tüm memeli benzeri sürüngenlerden tamamıyla ayrı bulundukları çok açıktır. Bu kanıt, yaratılış modelinin dayandırıldığı tahminle uyumludur ve evrim kuramıyla doğrudan bir zıtlık içermektedir.
Kemp senaryosuna göre, cynodont dışındaki memeli benzeri sürüngen seviyesinin ani kitlesel yok oluşlarından sonra, memeli benzeri sürüngenlerin üçüncü ve son evresi, hızlı bir yayılımdan kaynaklanmıştır. Bu yayılım da, Geç Permiyen kayalarının erken seviyelerinde bulunan cynodont’ların ortaya çıkmasına neden olmuştur. Kemp’in şu sözlerine göre, Cynodont’lar, ilk ortaya çıktıkları andan itibaren hep cynodont’lardır:
Bu ilk formlar birkaç açıdan Trias cynodont’larına göre daha ilkel olmalarına rağmen yine de şüphe götürmez bir şekilde evrimin cynodont seviyesindedirler.38
Sonuçta “gelişmiş” cynodontlardan birinin, memelilerin başlangıcı olduğuna inanılmaktadır. Fakat, hangisinin gerçek ataya en yakın konumda bulunduğu bir tartışma konusudur.
İki memeli benzeri sürüngenin, Morganucodon (Eozostrodon olarak da isimlendirilir) ve Kuehneotherium’un, sürüngenler ile memeliler arası en kesin geçiş formlarını temsil ettiği varsayılmaktadır. Morganucodon’un farklı bireylerini temsil eden binlerce fosil parçası bulunmuştur. Bulunan malzeme, Güney Afrika Karoo Süpergrubu’nun Kızıl Yatakları’nda bulunmuş olan iki benzer cinse ait yapılar, Çin’in Lufeng Kızıl Yatakları’nda bulunmuş olan çeneler ve bütün bir kafatası, Wales’te bulunmuş olan dişler, çeneler, kafatası parçaları ve postcranial iskeletlerden ibarettir.39 Wales’te çıkarılan Kuehneotherium’dan sadeceçene parçaları ve ayrık dişler bulunabilmiştir. Bu canlılar, yaklaşık on santim uzunluğunda oldukça küçük canlılardı. Bu bulguların tümü jeolojik sütunun Üst Trias döneminde yer almıştır.40
Bunlar, sürüngen biçimi çene ekleminin yanında memeli biçimi çene eklemine de sahip oldukları iddia edilen canlılardır. Memelilerde, alt çenenin her iki yarısında bulunan tek bir kemik vardır. Bu kemik, dişleri taşıdığı için dentary olarak isimlendirilir ve kafatasının squamosal bölgesiyle doğrudan bağlantılıdır. Sürüngenler, alt çenelerinin her iki yarısında altışar kemiğe sahiptirler. Çene kemiklerinden biri olan eklem kemiğinin (articular) memelilerde bulunmayan kuadratum kemiğiyle birleşmesi sonucu çene ile kafatasının bağlantısı dolaylı olmaktadır. Memeliler ile sürüngenler arasındaki bir diğer temel fark ise, ister canlı ister fosil, sürüngenlerin tümünde kulaklarda çubuk benzeri, columella olarak bilinen tek bir kemiğin bulunmasıdır. Memelilerin kulaklarında üç kemik bulunmaktadır: Çekiç, Örs ve Üzengi kemikleri. Evrimciler, üzengi kemiğinin columella’ya karşılık geldiğini ve sürüngenlerdeki kuadratum ile articular kemiklerin de sırasıyla memeli kulaklarındaki örs ve çekiç kemiklerinin yerine bir şekilde geçtiğini iddia etmektedirler. Fakat bu olaylar gerçekleşirken ara seviye formlarının duyma işini nasıl başardıkları konusunda hiçbir açıklama yapılmamaktadır.
Yukarıda açıklanan fikirdeki bir diğer zorluk, tek bir kulak kemiği ile çok sayıda çene kemiğine sahip binlerce sürüngen fosili ve üç kulak kemiği ile tek bir çene kemiğine sahip binlerce memeli fosili bulunurken, üç çene kemiği ile iki kulak kemiğine sahip olma gibi bir ara seviye form özelliği taşıyan tek bir canlı fosilinin bile bulunamamasıdır.
Morganucodon41 ve Kuehneotherium’un42 her ikisi de alt çenelerinde, sürüngen kemiklerine tam takım olarak sahiptiler. Üstelik bu canlılar, sürüngen çene eklemine ek olarak bir memeli (squamosal–dentary) çenesine de sahip oldukları ileri sürülen sürüngen–memeli arası ara seviyeler olarak düşünülmelerine rağmen, sahip oldukları sürüngen çene ekleminin (kuadratum–articular) işlevsel öneminde hiçbir azalma olmamıştır. Kermack ve diğerleri şöyle demektedir:
Morganucodon’un çenesindeki diğer kemiklerin en çarpıcı niteliği, cynodont’a özgü nitelik olmalarıdır. Cynognathus gibi tipik bir gelişmiş cynodont’a kıyasla, var olan diğer çene kemikleri, ne büyüklük ne de yapının karmaşıklığı açısından bir azalma göstermektedir. Morganucodon memelisindeki sürüngen çene eklemi, Cynognathus sürüngenindeki sürüngen çene eklemi kadar güçlüydü. Bu, oldukça beklenmedik bir şeydir.43
Bu yazarlar, evrimcilerin uzun zamandan beri, erken cynodont’ lardan geç cynodont’lara geçişte çene ekleminde ilerleyen bir zayıflamanın var olduğuna ve bu zayıflamanın, ilk memelilere kadar devam ettiğine inandıklarını anlatmaktadırlar (cynodont’lar, “gelişmiş” memeli benzeri sürüngenlerdi). Eğer memeliler sürüngenlerden evrimleşmişlerse ve sürüngen çene eklemi ile memeli çene eklemi arasında kademeli bir evrimsel yer değiştirme gerçekleşmişse, beklenilen şey böyle olurdu. Kermack ve çalışma arkadaşları şimdi bu fikirlerini reddetmektedirler. Çünkü, Cynognathus’un sürüngen çene eklemi çok güçlüdür ve Morganucodon’un alt çenesi de Cynognathus’unkine çok benzemektedir.
Bu nedenle Morganucodon’un, standart sürüngen biçimli, güçlü bir çene eklemine sahip olduğu konusunda hiçbir şüphe yoktur. Morganucodon’la ilgili olarak elimizde var olan bulguların hemen hemen hepsi, bir bütün halinde olmayan kemiklerden oluşmasına rağmen, kuadratumun hâlâ articular kemikle bağlantılı bir şeklinde olan bir çene parçası bulunmuştur. Bundan dolayı bu canlıda, bir sürüngen çene ekleminin var olduğuna dair hiçbir şüphe kalmamıştır. Fakat, bu sürüngen çene ekleminin yanında,Morganucodon ve Kuehneotherium, dentary ve squamosal kemikleriyle bir bağlantı noktasına sahip midir? Eğer öyleyse, bu, memeli biçimi çene ekleminin yeni başlamış oluşumunu mu göstermektedir?
Kermack ve meslektaşları, bunun, Morganucodon ve Kuehneotherium için, kanıtlanmış olduğuna kesinlikle inanmaktadırlar (ayrıca bunun, birçok farklı memeli benzeri sürüngen grubunda da aynı olduğu söylenmektedir).44 Bu inancın dayanağı nedir? Bu inanca ne kadar güçlü tutunulursa tutunulsun, bu inanç çıkarımlara dayanmaktadır. Kanıtlar son derece parça parçadır ve kafatasının squamosal kemiği ile dentary’nin bağlantılı bir halini gösteren hiçbir fosil yoktur. Gerçekte, bozulmamış, bütün halde tek bir alt çene bile bulunmamaktadır; bu tür modellerin tümü parçaların yeniden bir araya getirilip birleştirilmesiyle oluşturulmaktadır.
Bu canlılarda squamosal–dentary ekleminin bulunduğuna dair kanıt nedir? Bunun kanıtı dentary’de var olduğu iddia edilen bir kondildir. Kondil, bir diğer kemiğin fossa olarak isimlendirilen çukur parçasıyla birleştirilmesiyle yumrulu–yuvalı eklem oluşturan, yuvarlak bir kemik ucu çıkıntısıdır. Memelilerde, kafatasının squamosal kemiği ile bağlantılı halde olan dentary’nin arka ucunda çok belirgin bir kondil vardır. Squamosal kemik, kondil yerine geçebilecek bir fossadan oluşur ve buradaki bağlantı çene eklemini oluşturur. Morganucodon ve Kuehneotherium’da dentary, squamosal’a kadar ulaştığı inancını teşvik edecek kadar arka tarafa doğru yeterli biçimde uzama göstermiştir. Dentary üzerinde ileri sürülen bağlantı noktası için kondil adı kullanılmıştır.
Bu canlıların dentary kemiklerinin squamosal ile gerçek bir teması olup olmadığı sadece çıkarıma dayanmaktadır. Ayrıca dentary ve squamosal arasında gerçek bir temas olsa bile, bu durumda sürüngen çene ekleminin yanında bir memeli çene eklemi oluştuğunu söylemek mümkün müdür? Unutmamalıyız ki, bu canlılar tümüyle gelişmiş, güçlü bir sürüngen çene eklemine sahiptiler. Bu tür bir çene ekleminin gerektirdiği anatomi, yani kasların bağlantı biçimi, yerleşimi ve kan damarları ile sinirlerin yerleşimi, bir memeli çene ekleminin gerektirdiğinden çok daha farklı olmalıdır. Öyleyse, güçlü ve tümüyle işlevsel bir sürüngen çene eklemi, bir memeli eklemine nasıl uyum sağlayabilir?
Probainognathus ve Diarthrognathus’taki çift çene eklemiyle ilgili benzer iddiaların sorgulanması önemlidir. Probainognathus ve Diarthrognathus, memelilerin doğrudan kuramsal atalarına çok yakın olarak gösterilmişlerdir. Probainognathus ile ilgili olarak Kemp şöyle der:
Probainognathus’unmemelilerle akraba olduğunun çokça söylendiği ikinci özellik, squamosal ve dentary arasındaki ikincil bağlantıdır. Gerçekte, bu kemikler arasında gerçek bir bağlantı olup olmadığı biraz şüphelidir (Crompton ve Jenkins, 1979)...45
Diarthrognathus ile ilgili olarak Gow şöyle der:
Genellikle, Clarens Oluşumu’ndan (Cave Sandstone) (Crompton, 1958) Diarthrognathus adlı ictidosaur’un, memeliler ve cynodontlar arası ara seviye formlarının beklenen morfolojik derecelerini sergilediği varsayılmaktadır; ve özellikle, bu canlının memeli ve sürüngen çene eklemlerinin her ikisine de sahip olduğu düşünülmektedir. Ancak Crompton’un, alt çene morfolojisi ve onun kafatası ile bağlantısı konusunda yapmış olduğu yorumlar yanlıştı ve Crompton, hepsi olmasa da yorumlarının bazılarının yanlışlığını eserlerinde de dile getirmişti. (Crompton, 1972).46
Böylece görüyoruz ki, bu iki canlının hem memeli hem de sürüngen çene eklemlerine bir arada sahip oldukları fikrine evrim çevrelerinde karşı çıkılmıştır. Bu canlıların tümünün soyu tükenmiştir; geri kalan sadece, son derece parçalanmış fosillerdir. Bu canlıların parçalarının bir araya getirilmesi ve bunların işlevlerinin gözle canlandırılmasındaki tutum, ümit edilen önyargılı fikirlerden önemli oranda ve sıkça etkilenmiştir. Evrimciler, sürüngenlerin memelilere evrimleştiğinden emindirler. Bu, sürüngen çene ekleminin, memeli çene eklemi ile yer değiştirmesini gerektirecektir. Günümüze kadar gelen bulgular, aşırı derecede parçalanmış ve eksik olduğu için, insanların onlara baktıklarında, orada “görünen” şeyi, var olan olarak değil de görmek istedikleri bir şey olarak görmeleri mümkündür. Şüpheleri ortadan kaldıran bir sonuç olarak, güçlü, tümüyle işlevsel bir sürüngen çene eklemine sahip bir canlı ile, güçlü, işlevsel bir memeli çene eklemine sahip tek bir ara seviye canlısı bile bulunamamıştır. Morganucodon ve Kuehneotherium dahil olmak üzere tüm sürüngenler, çenelerinde, tam takım sürüngen kemiklerine ve tüm memeliler de, ister canlı ister fosil olsun, alt çenelerinin her iki tarafında tek bir kemiğe sahiptir. Hiçbir ara form bulunamamıştır.
Sürüngen Kulaklarına Karşı Memeli Kulakları
Çene eklemleriyle ilgili kanıtları, duyu organlarıyla ilgili kanıtlardan ayırmak mümkün değildir. Daha önceden de sözü edildiği gibi, evrimciler şuna inanmaktadırlar: Dentary dışındaki sürüngen çene kemikleri, çenedeki işlevleri yavaş yavaş azalırken, yeni işlevler gerçekleştirmeye evrimleşmekte ya da yok olmakta serbesttiler. Böylece, kuadratum ve articular kemikler serbest kaldılar (aslında, Morganucodon’da bu kemikler, dentary’e sıkıca tutturulmuşlardı) ve bir şekilde, orta kulağa doğru harekete geçtiler; ve sonuçta, sırasıyla örs ve çekiç kemiklerini oluşturdular. Bu, sürüngenlerdeki üzengi kemiğinin (columella) kulak zarıyla (tympanum) olan bağlantısının serbest hale gelmesini ve articular’ın retroarticular çıkıntısının kulak zarına bağlanmasını gerektirecekti (çünkü, sürüngendeki articular kemiğin, memelide kulak zarıyla bağlantı halinde olan çekiç kemiğine dönüştüğü varsayılmaktadır). Bunlar bir şekilde gerçekleşirken, sürüngen atadaki kuadratum kemiğinin serbest kalması, orta kulağa doğru hareket etmesi ve kendisini, üzengi kemiği ile çekiç kemiği arasında bir yere sığdırması gerekiyordu. Bu tür yer değiştirmeler olurken, tüm bu kemiklerin, tümüyle farklı işlevde duyu organları olabilmeleri için, bir şekilde mucizevi olarak yeniden biçimlenmeleri ve inşa edilmeleri gerekiyordu.
Böylesine inanılmaz bir senaryoyu destekleyecek bir fosil kanıtı kesinlikle bulunamamıştır. Kuehneotherium’un, Theria (keseli ve plasentalı) memelilere giden direkt yolun üzerinde bulunduğu varsayılmasına karşın kulak kemikçikleri yoktu. Kemp şöyle demektedir:
Theria’ların kulak kemikçiklerinin hangi evrede evrimleştiği bilinmemektedir. En erken ve ilkel Theria olan Kuehneotherium bu kemiklerden yoksun olmalıydı, çünkü artdentary kemiğini barındırabilmek için dentary’nin iç yüzünde hâlâ bir oluk bulunmaktadır.47
Bir sürüngenin bir memeliye olan evrimsel dönüşümünün bu hayatî evresinde, sürüngen çenesindeki articular ve kuadratum kemiklerinin yeni oluşmaya başlamış memelinin kulağına hareket ederek memeli kulağında örs ve çekiç şeklinde yeniden şekillenmesinin, fosil kaydında hiç belgelenmemesi tuhaf görünmektedir. Elbette ki, bu rastgele genetik hataların ve doğal seçilimin böylesine mucizevi organları inşa edebilmesi için gerekli olan milyonlarca yıl boyunca, yeterli bir fosil kaydı bırakılmış olmalıydı.
Böyle varsayılan bir süreçle ilgili anatomik sorunlar, güçlü ve etkili bir çene ekleminde işlev görecek şekilde tam ayarlanmış iki kemiğin nasıl kendilerini çıkarıp orta kulağa sokabileceğini, nasıl tümüyle farklı duyu organında yeniden şekillenmiş üzengi kemiği ile birlikte işlev görecek şekilde örs ve çekiç kemiklerine dönüşeceğini ve bu olaylar devam ederken canlının, çiğnemeye ve duymaya nasıl devam edeceğini hayal etmekten çok daha büyüktür! Böylesine aşılması güç bu sorun, hiçbir sürüngende bulunmayan ve nereden geldiği konusunda ipucu bile verebilecek hiçbir kanıtı olmayan bir organ olan Corti organının memelilerde esas duyu organı olduğunu düşündüğümüz zaman, önemini kaybetmektedir.
Corti organı, son derece karmaşık bir organdır. Okuyucuların, açıklama için, anatomi konusunda standart bir kitaba başvurmalarını öneriyoruz. Bir kişinin bu olağanüstü biçimde tasarlanmış organın karmaşıklığı karşısında hayran kalmaması mümkün değildir. Bu organın sürüngenlerde bir benzeri yoktur. Sürüngenlerde, bu organın türeyebileceği tipte olası bir yapı da yoktur. Bu organ, tümüyle alışılmadık ve yeni bir organ olduğu için, yoktanyaratılmış bir organ olmak zorundadır.
Evrim kuramına göre, evrimsel değişimlerin tümü genlerin yeniden üretilmesi sırasında gerçekleşen hataların bir sonucu olarak ortaya çıkmaktadır. Devam eden mutasyonların ortaya çıkardığı her değişim, kendisinden önceki formlardan üstün olmalıdır. Eğer evrim gerçekse, Corti organının, kendisine uygun bir şekilde yeniden inşa edildiği ve çene kemiğinden iki tane kemiğin sürüklenip tekrar tasarlandığı kulakta işlev görmesi için, olağanüstü bir şekilde uyarlanmış binlerce hata dizisi sonucu kademeli olarak yaratıldığına inanmamız gerekir. Üstelik, her ara aşamanın tümüyle işlevsel olmakla kalmayıp, aynı zamanda kendinden önceki aşamalardan daha üstün nitelikler de taşıması gerekmekteydi. Ve bütün bunlar başarıyla sonuçlandıktan sonra bile, bugün hâlâ, memeli duyu organları kadar etkin çalışan eski yapıda duyu organlarına sahip sürüngenler ve kuşlar vardır.
Gerekli Olan Diğer Değişimler
Üstelik, yukarıda anlatılan olağanüstü değişimlerin hepsi gerçekleşirken bu canlılar, (genetik hatalar yoluyla) yeni bir üreme biçimi, meme bezleri, sıcaklık ayarı, saç ve yeni bir nefes alma biçiminin oluşumunu içeren olağanüstü yeni fizyolojik ve anatomik organlar ve süreçler icat etmişlerdir.
Memelinin göğüs kemeri yapısı, temel anlamda bir sürüngeninkinden farklıdır. Sürüngende bu kemik, korakoit (coracoid) kemikler yoluyla göğüs kemiğine bağlanır ve göğsün bir parçasını oluşturur. Bu durum, memelilerde böyle değildir. Sürüngenlerde göğsün ön parçası serttir ve genişleme yeteneği yoktur. Memelilerdeki göğüs, genişleme özelliğine sahiptir. Memelilerde göğüs ve karın boşlukları, lifli kaslardan oluşan bir organ olan diyaframla birbirlerinden ayrılmışlardır. Sürüngenlerde diyafram olmadığı için, göğüsleri kapalı bir kutu değildir. Yukarıda anlatılanların bir sonucu olarak sürüngenler, memelilerin yaptığı gibi nefes alamazlar. Göğüsleri memelilerde olduğu gibi dönüşümlü olarak genişleyip daralmaz. Sürüngenler, ağızlarıyla nefes almalıdırlar.
Sürüngenlerde, memeli diyaframına benzer ya da eş herhangi bir yapı yoktur. Sürüngenlerde, diyaframın türeyebileceği herhangi bir yapı da bulunmamaktadır. Yine, varsayılan sürüngen atanın farklı bir yöntemle başardığı bu işlevin yerine getirilebilmesi için, karmaşık bir yapının (bir dizi hata yoluyla) yoktan yaratılması gerekirdi.
Memeli Evrimindeki Büyük Boşluk
Memeli benzeri sürüngenler, varsayımsal olarak, sürüngenlerin başlangıcında ortaya çıkmışlar ve Permiyen ile Trias süresince kademeli olarak daha da çok memeli benzeri sürüngenler halini almışlardır. Sonuçta, Trias sonunda, ilk gerçek memelilerin görünmesiyle doruk noktasına ulaşmışlardır. Memeli benzeri sürüngenler, daha önceden tüm dünyaya yayılmış ve tüm sürüngenlerden sayısı en fazla olanlara ait olmalarına rağmen, bu zamanın sonunda neredeyse soyları tükenmiştir. Yüksek oranlarda uyum sağlayan canlıların çok sayıda ürediği ve bu canlıların, şartlara daha az uyum gösterenlerin yerini aldığını varsayan doğal seçilim aracılığıyla evrim oluştuğunu iddia eden evrim kuramına göre, artık galip çıkan memelilerin çok büyük rakamlara çoğalmaları ve dünyaya egemen olmaları beklenmektedir. Ancak, çok garip bir şey olmuştur. Net söylemek gerekirse memeliler, bundan sonraki 120 milyon yıl boyunca sahneden silindiler! Varsayılan bu uzun zaman süreci boyunca dinozorları, karada yaşayan başka birçok canlıyı, deniz sürüngenleri ve uçan sürüngenleri kapsayan “sürüngen benzeri” sürüngenler, yeryüzünü dolaştılar. Bununla beraber, memeli benzeri sürüngenlerin yerini alan “en çok uyum sağlayan” memeliler neredeyse hiçbir yerde bulunamadılar. Sözde 120 milyon yıldan daha uzun bir zamanı kapsayan Jura ve Tebeşir dönemlerine ait memeli fosil kalıntılarının çoğu iki avuca sığacak kadardır. Bu memelilerin çoğu, birkaç diş aracılığıyla bilinmektedir. Eğer evrim en uygun olanların hayatta kalmasını kapsıyorsa ve en uygun olanlar, çok büyük rakamlara kadar üreyenler olarak tanımlanıyorsa, memelilerin kökenini gerçekten de çok garip bir şey teşkil etmektedir. Memeliler, çok az sayılarda hayatta kalabildikleri için, evrim, uyum sağlayamadan hayatta kalanlar üzerinde gerçekleşmiş görünmektedir!
Evrimciler bizim, memeli evriminin 120 milyon yıl boyunca durgun kaldığına inanmamızı istemektedirler. Evrim kuramına göre, 120 milyon yıl boyunca görünüşte çok az sayıda olan memeliler, küçük ve özelleşmemiş formlarından ötürü evrimleri durmuş bir halde kaldılar. Daha sonra, dinozorları da kapsayan sürüngenlerin çoğu, jeolojik zamana göre göz kırpması kadar kısa bir sürede yok oldular ve maymunlar, balinalar, yarasalar, kemirgenler, tek toynaklılar, çift toynaklılar gibi canlıların oluşturduğu otuz iki memeli sınıfı aniden, tümüyle gelişmiş biçimde ortaya çıktı.
Yaratılışa olan eğilimimizden dolayı durumu abarttığımız düşünülüyorsa, o zaman dünyanın önde gelen evrimcilerinden biri olan George Gaylord Simpson’un bu konudaki yorumlarına bir göz atalım. Gaylord şöyle demiştir:
Dünyadaki yaşam tarihine bakıldığında en şaşırtıcı olay, Sürüngenler Çağı olan Mezozoik’ten, Memeliler Çağı’na olan değişimdir. Sanki, tüm başrollerin sürüngenler ve özellikle de çok sayıda ve şaşırtıcı çeşitlilikte olan dinozorlar tarafından paylaşıldığı bir sahnede, perde aniden aşağı inmekte ve hemen tekrar yukarıya kaldırıldığında dekor aynı fakat dinozorların hiç görünmediği, diğer sürüngenlerin önemsiz rolleri üstlendiği ve önceki sahnede çok az görünen memeli çeşitlerinin tüm başrolleri oynadığı yepyeni bir oyuncu kadrosu sahneye çıkmaktadır.48
Yeniden vurgulamalıyız ki, önceki sahneler, evrim zamanına göre 120 milyon yıl sürmüştür. Bir evrimci, sanki burada ciddi bir problem yokmuş gibi rol yapmaya kalkarsa, Simpson’un bu olaydan, dünya tarihindeki en şaşırtıcı olay olarak söz ettiğini hatırlasın. Evrim varsayımı atılıp kökenlerle ilgili yaratılış modeli kabul edilirse, elbette ki bu sorun kendiliğinden ortadan kalkacaktır. Ayrıca, Simpson’un, bu sorunu “yaşam tarihindeki en şaşırtıcı olay” olarak tanımlarken, karmaşık omurgasızların Kambriyen kayaları arasında aniden, tümüyle gelişmiş biçimde ortaya çıkışlarını “yaşam tarihinin en büyük sırrı” olarak tanımladığını da hatırlamaktayız. Evrimciler gerçekte, yaşam tarihindeki birçok olayı şaşırtıcı ve açıklanamaz olarak yorumlamak zorunda kalmaktadır.
Evrimcilerin 100 Milyon Yıllık Şoku
Son yıllarda yapılan bir keşif, evrimcilerin yeni bir şaşırtıcı sürprizle karşılaşmalarını sağladı. Evrimciler daima memeli benzeri sürüngenlerin, memelilerin evrimleştiğini varsaydıkları Geç Trias’tan sonra geldiğine inandıkları Orta Jura olarak adlandırılan dönemde nesli tükendiğini varsaymaktadırlar. Evrimciler, memeli benzeri sürüngenlerin memelilere dönüştüğü ve paleontologlar Orta Jura kayalarından daha genç kayalarda bunları bulamadığı ya da en azından onların kimliklerini belirleyemediği için memeli benzeri sürüngenlerin soylarının 160 milyon yıl önce tükendiğini düşünmüşlerdir. Pek çok evrimci paleontolog ve biyolog, Fox, Youzuryshun ve Krause’nin, Kanada’nın Alberta eyaletindeki Geç Paleosen kayaları arasında yaklaşık 60 milyon yıl yaşında memeli benzeri bir sürüngene ait fosil buldukları iddialarına inanmayı güç bulmuşlardır.49 Orta Jura’dan Geç Paleosen’e kadar olan zaman aralığının, 100 milyon yıllık bir süreyi kapsadığı varsayılmaktadır. Fox, Youzuryshun ve Krause, bu memeli benzeri sürüngeni, Chronoperates paradoxus (chronos: zaman; perates: gezen; paradoxus: beklentilere zıt) olarak isimlendirdiler. Fox ve çalışma arkadaşları, buldukları fosili memeli benzeri sürüngen olarak belirleyen inandırıcı kanıtlar sunmaktadırlar. Pennsylvania Üniversitesi’nden Neil Shubin şöyle demiştir: “Eğer Chronoperates, Trias’ta bulunsaydı, Fox’un bu fosili bir memeli benzeri sürüngen olarak belirlemesinde hiçbir sorun olmazdı.”50
Bu gerçekler evrimciler için, can sıkıcı sorular yaratmaktadır. Eğer memeliler, varsayıldığı gibi, doğal seçilim yoluyla memeli benzeri sürüngenlerden evrimleşmiş ve onların yerini almışlarsa, memeli benzeri sürüngenler bundan sonraki 100 milyon yıl boyunca nasıl memelilerle birlikte yan yana hayatta kalabilmişlerdir? Eğer Chronoperates (ya da onun memeli benzeri sürüngen ataları) Orta Jura ve Geç Paleosen arasındaki 100 milyon yıllık süre içerisinde hayatta kalabilmişse, neden bunların fosilleri inanılmaz derece nadir bulunmaktadır; ki bu güne kadar tek bir fosil bulunabilmiştir. Bu 100 milyon yıl içerisinde, her yıl yalnızca bir milyon memeli benzeri sürüngenin yaşadığı ve öldüğü düşünülürse, bu zaman sürecinin bütününde 100 trilyon memeli benzeri sürüngen yaşayıp ölmüş olacaktır; ama şimdiye kadar yalnızca tek bir fosil bulunabilmiştir. Bu durum, Afrika kıyılarına yakın yerlerdeki derin sularda büyüyüp geliştikleri keşfedilinceye kadar, coelacanth balıklarının, bundan 70 milyon yıl önce soylarının tükendiğine inanıldığı duruma benzemektedir. Evrim senaryosunda, ciddi bir yanlışlık olduğu görülmektedir. Bu gerçekler, evrimsel zaman çizelgesinde ve kuramsal süreçlerde önemli şüpheler ortaya koymaktadır.
Memelilerdeki Büyük Çeşitlilik
Diğer başlıca hayvan ve bitki gruplarında olduğu gibi, memeliler de daima büyük farklılık gösteren bir grup olmuştur. Bugün, yaklaşık 4300 farklı memeli türü bilinmektedir ve binlerce türün de soyu tükenmiştir. Son birkaç heyecan verici keşif, memeliler hakkında bildiklerimizi şüphesiz genişletecektir. En heyecan verici ve şüphesiz yararlı keşiflerden biri, Amerikalı ve Rus bilim adamlarından oluşan bir ekip tarafından Gobi Çölü’nde gerçekleştirildi.51 Orada bu ekip, içerisinde barındırdığı dinozor fosilleri ile ünlü olan Flaming Kayalıkları bölgesinde memeli fosillerden oluşan zengin bir kaynak buldu. Ekip, şimdiye kadar “mükemmel derecede korunmuş” 187 adet memeli kalıntısı ortaya çıkardı; bunların pek çoğu, tüm iskeletten oluşuyordu. Amerikan ekibinin liderlerinden biri ve Amerikan Doğa Tarihi Müzesi’nde kıdemli bir bilim adamı olan Michael Novacek, bulunan kalıntıların, ilk bulguların bulunduğu tarih olan 1922’den bu yana Gobi Çölü’nden getirilen tüm memeli fosillerinden sayıca fazla olduğunu bildirdi. Son yıllarda yapılan toplantıda bulguları görme fırsatı yakalayan araştırmacılar, “fosillerin ne kadar sağlam ve tüm halde olduğunu gördüklerinde hayrete düştüler.” Bu fosiller, Tebeşir kayaları arasından çıkarılmışlardı ve evrimciler, fosillerin temsil ettiği memelilerin, dinozorların yok oluşlarından yaklaşık on beş milyon yıl önce, yani bugünden yaklaşık seksen milyon yıl önce var olduklarına inanmaktadırlar.
Bu keşifler evrimcileri, memelileri betimleme biçimlerini yeniden gözden geçirmeye zorlamıştı; çünkü bu bulgular, memelilerin, dinozorların yok oluşlarından önce çok yaygın ve çeşitli olduklarını açığa çıkarmıştır; Novacek bunların, dinozorlara doğrudan bir rakip olmadıklarını söylemiştir. Gobi Çölü bölgesinde, dinozor, memeli, kertenkele, timsah ve kaplumbağa fosilleri bulunmuş ve sonuçta ortaya çıkan eski yaşam tablosu, gittikçe daha çok yaratılışçıların öngördükleri tabloya benzer bir tablo olmuştur.
Özellikle heyecan verici olan keşif de, bozulmamış orta kulak kemikçikleri ve diğer önemli kafatası özelliklerini içeren memeli kafatası fosillerinin keşfidir. Yaratılışçılar, orada bulunan şeylerin her türünün, hiçbir geçiş özelliği kanıtı olmaksızın tümüyle bozulmamış olmasını öngörmekte ve bulguların, bilim dergilerinde yayınlanmasını sabırsızlıkla beklemektedirler. Şimdiye kadar sadece kısa ve sözlü raporlar bulunmaktadır. Bu durumda eğer evrim gerçekse, memelilerin erken Üçüncü Zaman (Tersiyer) kayaları arasında görünen “nefes kesici evrimsel yayılımı”na (evrimcilerin dediği gibi) neden olan, evrimleşen ilkel memeli formlarının birçok ara formunu bulmamız gerekmektedir.
Bir diğer dikkate değer memeli fosili bulgusu, Almanya’nın Frankfurt şehrinden 19 km uzakta bulunan Messel’in bir maden ocağında bulunmuştur. Bu maden ocağı 1986’da Hesse Eyaleti tarafından doğal korunma alanı olarak ilan edilmiştir. Bu yerden şimdiye kadar, yaklaşık otuz iki memeli takımının on dördüne ait kırktan fazla memeli türü örneği çıkarılmıştır.52 Bu fosillerin, bugünkü evrimsel zaman ölçeğinde yaklaşık 50-35 milyon yıl önceye, Eosen Çağı’na ait oldukları saptanmıştır. Bu fosiller olağanüstü derecede iyi korunmuştur ki, birçoğu yumuşak vücut taslağı temel alınarak yeniden yapılandırılmıştır. Pek çoğunun mide içerikleri bozulmamıştı, örneğin bir yarasanın bağırsağında alacakaranlıkta ya da gece uçan güveler bulunmuştur. Bu örnek de, bu yarasanın geceleri beslendiğini ve pek çok günümüz yarasasında bulunan ekolokasyon (sesin yankılanmasından faydalanarak, bir cismin bulunduğu yön ve uzaklığı saptama) organına sahip olduğunu göstermektedir. Buradan çıkarılan fosillerin içinde, tek ve çift toynaklılar, lemur benzeri yarı maymunlar, kirpi benzeri böcekçiller, kemirgenler ve keseli sıçan benzeri keseliler vardı. Ayrıca burada, Edentata takımına ait karıncayiyenlerden Eurotamandua joresi’nin de bir fosili bulunmuştur. Bu fosil sözde 50 milyon yıl yaşında olmasına rağmen, günümüz karıncayiyen cinslerinde bulunan oldukça karmaşık belirleyici özelliklerin tümüne sahipti. Ayrıca, en eski pangolin olarak bilinen Eomanis waldi’nin de bir fosili bulundu. Pangolinler, üst üste binen geniş pullardan oluşan bir örtüye sahip karıncayiyenlerdir. Bulunan E. waldi, yine sözde 50 milyon yıl yaşında olmasına rağmen, modern pangolin türlerine benziyordu.
Gerhard Storch bu çok önemli fosil mezarlığı hakkındaki raporunda,53 burada fosilleri bulunan tüm memelilerin, başka kıtalardan Avrupa’ya göç etmiş olduklarını söylemek zorunda kalmıştır; çünkü, bunlar, hiçbir evrimsel tarih olmaksızın tümüyle gelişmiş biçimde ortaya çıkmışlardı. Storch’un memelilerin geldiği kıtanın Afrika olduğunu düşünmesi sadece bir çıkarımdır, çünkü Afrika’da böyle bir senaryoyu destekleyebilecek hiçbir fosil kanıtına rastlanmamıştır. Evrimciler, belirli bölgelerde canlıların aniden ve tümüyle gelişmiş biçimde ortaya çıkmalarını açıklayabilmek için göç fikrine sık sık başvurmaktadırlar. Tabi ki varsayılan bu göçün kaynağı olan bölgede, gerekli olan atalar da bulunmamaktadır.
Ana Memeli Grupları
Taksonomistler arasında ana memeli grupları konusunda bazı kararsızlıktan kaynaklanan görüş ayrılıkları vardır. Bazıları, günümüz memelilerini iki ana gruba ayırmaktadır. Böylece Mammalia Sınıfı’nın yaşayan memelileri, iki altsınıfa ayrılmaktadır: tekdeliklileri içeren Prototheria ve Theria altsınıfları. Bazı kişiler Theria altsınıfını da kendi içinde tekrar Metatheria ya da keseliler ve Eutheria ya da gerçek plasentalı memeliler olarak iki infra–sınıfa ayırmaktadır. Monotremata takımına ait günümüz tekdeliklileri, ördek gagalı ornitorenkleri, Ornithorhyncus’u ve Avustralya dikenli karıncayiyeni Tachyglossus’u ve Yeni Gine dikenli karıncayiyeni Zaglossus’u kapsar. Bu canlıların her ikisinin de hem çok ilkel, hem de ileri derecede özelleşmiş oldukları düşünülmektedir. Bu canlılar memelidirler; çünkü, yavrularını emzirmek için meme bezlerine sahiptirler, sıcak kanlıdırlar, kılları vardır ve memelilerin diğer niteliklerine de sahiptirler. Diğer yandan bir ornitorenk, bir ördek gagasına, perdeli ayaklara ve özellikle kuşlarda olduğu düşünülen yumurtlama özelliğine sahiptir; ve aynı zamanda bir sürüngen omuz kemerine ve birkaç sürüngen niteliği daha taşıyan postcranial bir iskelete sahiptir. Avustralya dikenli karıncayiyeni (echidna) ve ornitorenk’in kafatasları, yaşayan ve fosil halde bulunan diğer tüm memelilerden farklı olacak biçimde, ileri derecede özeldir. Romer bunları, yaşayan omurgalıların en tuhaf ve en yanıltıcı olanları arasında bulunan iki canlı olarak tanımlamıştır. Bunlar evrimcilerin var olmalarını hiç istemedikleri canlılardır. Memeli, kuş ve sürüngen niteliklerini (hiç birisi ara seviyede olmayan bu nitelikleri) taşıyan bu canlılar, bu üç omurgalı sınıfının ne atası ne de soyu olmayacaklardır. Tekdeliklilerin soy oluşu, evrimciler arasında bir tartışma konusudur. Bu canlıların fosil kaydı son derece yetersizdir ve Avustralya ile sınırlıdır. Bunların, erken bir evrede diğer memelilerden ayrıldıkları ileri sürülmektedir; fakat, bu tuhaf canlıların evrimsel kökenlerini belgeleyebilecek hiçbir geçiş formu bulunmamaktadır.
Keseliler (Marsupialia Takımı) göz önüne alındığında akla hemen üzerinde çok çeşitli keselilerin yaşaması nedeniyle Avustralya gelir. Ancak geçmişte keseliler Avustralya’dan çok Kuzey ve Güney Amerika’da ve hatta Avrupa’da çok daha yaygın halde bulunuyorlardı. Bu grubun en eski üyeleri oldukları varsayılan canlı fosilleri (yaklaşık seksen milyon yıl yaşında) Kuzey Amerika’nın geç Tebeşir kayaları arasında bulunmuştur. Bu canlılar, yaşayan keseli sıçanlara çok benziyorlardı. Hemen hemen tüm keseliler (fakat tümü değil), tam olgunlaşmadan doğan yavruların memeye yapışık bir şekilde gelişimlerini tamamlayabilecekleri barınak konumundaki bir keseye sahiptirler.
Keselilerin Güney Amerika’da verimli olarak çoğalmalarının Tersiyer Dönemi’nde olduğuna inanılmaktadır. Çoğu etçil olan bu canlıların büyüklüğü, keseli sıçandan ayıya kadar değişiyordu. Bu etçil keselilerden biri, büyüklük olarak jaguara, dişleri de süvari kılıcı dişli kaplanınkine benzeyen Thylacosmilus’tur. Antarktika’da bir keseli fosili Antarctodolops’un bulunması, keselilerin Antarktika’dan Avustralya’ya, bu iki büyük kara parçasının sözde birleşik olduğu zamanlarda göç etmiş olabileceği iddiasını destekliyordu.
Bugün Avustralya’da çok çeşitli keseliler yaşamaktadır. Bunlar, bildiğimiz kanguruların yanı sıra, bandicoot, Tazmanya “kurdu”, Tazmanya şeytanı, koalalar, vombatlar (içine toprak girmesini önlemek için kesesi geriye doğru açılan, çukur kazan bir hayvan) ile keseli “fareler,” “köstebekler,” “kediler” ve “sincaplar”dır. Avustralya’nın Pleistosen çökelimlerinden çıkarılmış birçok soyu tükenmiş büyük keseli bilinmektedir. Bunlar arasında, şimdiye kadar bilinen en büyük keseli olan, yaklaşık büyük bir gergedan kadar büyük Diprotodont ve yaklaşık bir aslan büyüklüğündeki Thylacoleo da vardır.
Evrimciler uzun zamandan beri, Avustralya keselilerinin büyük çeşidinin ata ya da atalarının, Güney Amerika’da birlikte yaşadıkları plasentalı memeliler Avustralya’yı istila edemeden, muhtemelen Antarktika yoluyla Avustralya’ya girdiklerini desteklemektedirler. Keseli memeliler artık plasentalı memelilerin rekabetinden kurtularak, sözde çeşitlenebildiler ve plasentalıların diğer bölgelerde yaptıklarını üstlendiler. Ancak bu, en olanaksız senaryodur. İlk önce, yukarıda anlatıldığı gibi, keseliler Antarktika yoluyla Avustralya’ya gitmenin bir yolunu buldukları sırada, plasentalılar Güney Amerika’da keselilerle birlikte, yan yana yaşıyorlardı. Eğer keseliler Avustralya’ya giden yolu bulabilmişlerse, plasentalıların da bu yolu bulmalarına ve belki de keselilerin önünden gitmelerine engel olacak hiçbir sebep olmayacaktır. Üstelik, eğer son yıllarda yayınlanan rapor doğru ise, görünüşe göre plasentalılar, memelilerle aynı zamanda Avustralya’ya varmışlardır. 1992 yılına ait bir gazetede Godhelp ve diğerleri Avustralya’nın Queensland eyaletinin güneydoğusundaki Mugon’a yakın bir yerde, tatlı su killeri içinde uçamayan bir plasentalı memeli fosili bulunduğunu bildirmişlerdir.54 Onlar, bu kayaların Erken Tersiyer Dönemi’ne ait, yaklaşık 55 milyon yıl yaşında olduğunu iddia etmekte ve şöyle demektedirler:
Avustralya’nın Erken Tersiyer Dönemi’ne ait uçamayan plasentalıların bulunması, plasentalıların Avustralya’ya Geç Tersiyer’den önce gelmemelerinden ötürü keselilerin Avustralya’nın Theria topluluklarında baskın bir rol oynadıklarına dair yaygın bir varsayıma karşı durmaktadır.
Daha sonra şöyle demektedirler:
Tingamarra porterorum, Avustralya’daki uçamayan plasentalı memelilerin en erkeni olarak bilinmektedir. Bu canlının Tingamarra Yerel Fauna’sındaki varlığı göstermektedir ki, Üçüncü (Senozoik) Zamanda Avustralya’da keselilerin egemen olmaları, bu tür plasentalıların bu kıtaya Erken Tersiyer Dönemi içinde varamayışlarına bağlanmamalıdır. Güney Amerika ve muhtemelen Antarktika’da olduğu gibi, plasentalılar ve keselilerin her ikisi de Senozoik Zamanın başında vardı. Fakat, bazı beklenmedik nedenlerden dolayı erken plasentalılar, Avustralya’da hayatta kalamadılar.
Bu fosiller için iddia edilen yaş, bundan yaklaşık 25 milyon yıl önce Geç Oligosen Çağı’nda yaşamış Avustralya erken keseli fosil bulguları için önerilen yaşın iki katıdır. Oligosen Çağı’na kadar, hiçbir geçiş formu izi olmaksızın tümüyle gelişmiş biçimde ortaya çıkmalarından ötürü, evrimciler, bu grupların çoğunun önceden farklılaştığını farz etmektedirler.
Godhelp ve diğerlerinin raporu yayınlanmadan önce, 1988’de yayınladığı kitabında Carroll şöyle demektedir:
Avustralya bölgesindeki keseli memelilerin fosil kaydı Geç Oligosen’de başlamaktadır... Bu zamana kadar, başlıca grupların pek çoğu zaten farklılaşmıştır. Keselilerin Avustralya’ya ilk ne zaman girdiklerini belgeleyecek doğrudan hiçbir kanıt bulunmamaktadır.
Keselilerin güney kıtalardaki kökeni ve dağılma yönü halen tartışılmaya devam edilen bir konudur... Marshall henüz hiçbir kesin kanıt olmadığını vurgulamaktadır.55
Keselilerin Kökeni
Bu, tahmin niteliğinde bir olay olmasına karşın evrimciler ve yaratılışçılar keseli memelilerin muhtemelen Güney Amerika’dan Antarktika yoluyla, karasal köprü ve bağlantıları kullanıp göç ederek Avustralya’ya ulaştıkları konusunda aynı fikirdedirler. Ama bu fikir birliği sadece buraya kadardır. Evrimciler, Avustralya’da bulunan keselilerin, göçmen keseli ataları Avustralya’ya ulaştıktan sonra çok farklılaştıklarına ya da evrimleştiklerine inanmaktadırlar. Diğer yandan yaratılışçı bilim adamları, bu canlıların her ana türünün ayrı ayrı yaratıldığına ve Avustralya’ya göç etmeden önce de aynı yapıda var olduklarına inanmaktadırlar. Fosil kaydı da kesinlikle yaratılış görüşünü desteklemektedirler. Avustralya’da yaşayan veya fosil halde bulunan keseliler arasında her büyüklükte, şekilde ve çeşitte keseli bulunmuştur. Bazıları fare kadar küçüktü. Bazıları ise bir gergedandan daha büyüktü. Fareye, zerdevaya, kurda, köstebeğe, kediye ve sincaba benzeyenler vardı. Bunların yanında, koalalar, bandicoot’lar, vombatlar ve kanguruları da içeren eşsiz keseli grupları da vardı. Eğer bu canlılar Avustralya’daki bir veya birkaç ortak atadan evrimleşmişlerse, sözde son elli altmış milyon yılı kapsayan fosil kaydı, bu büyük sayıdaki benzersiz canlıların ortak ata(ların)dan evrimleştiğini belgelemeye yetecek geçiş formları sağlamalıdır. Kanguruların, bandicoot’ların, vombatların, Tazmanya kurdunun, Tazmanya şeytanının, kuskusgillerin (philanger), dev Diprotodont’un, koalaların ve keseli “kediler,” “köstebekler” ve “fareler”in evrimsel kökenlerini belgeleyen atalara ve geçiş formlarına sahip olmamız gerekmektedir. Çok olmasa da elimizde fosiller bulunmaktadır ve bunların çoğu da temelde günümüz familyalarıyla aynıdır. Ancak, bu canlıların tek bir tanesinin bile evrimsel kökenini belirleyecek geçiş formlarına sahip değiliz. Bilimsel kanıtlar açısından, koalalar daima koalalar, kangurular daima kangurular, vombatlar da daima ilk ortaya çıktıklarından beri vombatlardı. Evrimciler bugün, hem plasentalıların hem de keselilerin birbirlerinden daha yaşlı olmadıklarına inanmaktadırlar ve keseliler hiçbir şekilde plasentalılardan morfolojik ve fizyolojik anlamda daha ilkel değildirler.56 Bu durumda tüm fosil kaydında olduğu gibi, kayalar yaratılış için güçlü olumlu kanıtlar sunmaktadırlar.
Plasentalı ve Keseli Memeliler Arasındaki Şaşırtıcı Benzerlikler
Kurama göre tüm evrim, sonuç olarak rastgele genetik hatalar, yani mutasyonlarla oluşmaktadır. Daha önceden de belirtildiği gibi, genellikle iddia edilen şey, karmaşık bir yapının ya da organizmanın, bir kereden daha fazla bağımsız biçimde evrimleşmesinin neredeyse imkansız olduğudur. Zaten karmaşık bir organ ya da organizmanın binlerce kuramsal ve rastlantısal “iyi” mutasyonların birleşimi sonucu evrimleşebileceğini hayal etmek yeterince zordur, hatta bunun ikinci kez olmasını hayal etmek zorun da ötesindedir.
Evrimciler, plasentalı ve keseli memelilerin, memeli benzeri ya da ilkel memeli ortak atadan ayrılarak bağımsız evrimsel yollar izlediklerine inanmaktadırlar. Yine evrimciler, özelleşmiş plasentalı ve keseli memelilerin o zamandan beri bağımsız olarak evrimleştiklerine inanmaktadırlar. Evrimsel süreç için, biri plasentalı, diğeri keseli birbirine çok benzeyen iki canlıyı ortaya çıkarmak, inanılmaz derecede zor ya da imkansız olacaktır. Bu canlıların her birinin evrimi, dünyanın farklı bölgelerinde avlanma olanağı ve yırtıcı hayvanlar tarafından avlanma tehlikesi olan farklı çevresel şartlar ve ortamlarda gerçekleşmiş olacaktır. Ama, hem fosil kaydında hem de bugün yaşayan keselilerde bu canlıların taklit ettiği çok çeşitli plasentalılar bulunmaktadır.
Güney Amerika’nın alt Miyosen kayaları arasında bulunan Necrolestes keselisi, plasentalı böcekçillere o kadar çok benzemektedir ki, uzun zaman boyunca plasentalı bir böcekçil olarak yanlış değerlendirilmiştir.57 Tazmanya “kurdu”, plasentalı kurtlara çok benzer. Carroll, bir Tazmanya “kurdu” olan Thylacinus’la ilgili olarak şöyle demektedir:
Diş ayrıntıları ile birlikte genel vücut biçimleri de plasentalı köpekgillerle çarpıcı derecede yakın bir benzerlik göstermektedir.58
Numbat ya da Avustralya karıncayiyeni olarak bilinen Myrmecobius, plasentalı karıncayiyenlerinkine çok benzeyen uzun bir buruna sahipti. Ve tabi ki, fare benzeri, kedi benzeri, köstebek benzeri Avustralya keselileri de bulunmaktadır.
Evrimciler, plasentalı ve keseli memelilerin, bundan yaklaşık 70-80 milyon yıl önce, geç Tebeşir’in bir döneminde ortak bir atadan ayrıldıklarına inanmaktadırlar. O zamanda ne plasentalı kurtların ne de Tazmanya yani keseli kurtların ataları, kurtlara ait tek bir özelliğe bile sahip değildi. Daha sonra dünyanın farklı bölgelerinde, önemli derecede ya da tümüyle farklı olması gereken ekolojik, biyolojik ve jeolojik şartlar eşliğindeki rastlantısal genetik hatalar sonucu, amaçsız bir evrim süreci yoluyla, diş ayrıntıları dahi birbirine çok benzeyen iki canlı yaratıldı. Ve benzer olaylar bir kez değil, birçok kez yaşandı. Ne kadar inanılmaz bir inanç!
Özelleşmiş Plasentalı Memelilerin Kökeni
Temel bir yaratığın kendisiyle bazı nitelikleri paylaşan ayrı bir canlıya dönüşümünü belirtmek başka, eşsiz bir yaşam tarzı için tasarlanmış ve böylece çok özel niteliklerle donatılmış canlıların kökenini belgelemeye çalışmak ise başkadır ve çok daha belirleyicidir. Böyle özelleşmiş özelliklerden bazıları şunlardır: uçan böceklerin, uçan sürüngenlerin ve uçan memelilerin kanatları; kuşların tüyleri; kuşlardaki, dinozorlardaki ve ornitorenkteki ördek gagası; yarasalardaki ekolokasyon organı; memelilerdeki kulak kemikçikleri; balinaların büyük derinliklere dalabilmek için ihtiyaç duyduğu kafatası özellikleri; daha önceden anlatılan farklı dinozorların pek çok benzersiz özelliği ve daha birçokları. GEÇİŞ FORMLARININ EN AÇIK VE EN KOLAY SAPTANABİLECEĞİ YER BURASIDIR. GEREKLİ OLAN GEÇİŞ FORMLARININ EN ÇOK İHTİYAÇ DUYULDUĞU YER BURASIDIR. EVRİM TEORİSİNİN DEĞİŞMEKSİZİN YETERSİZ KALDIĞI YER BURASIDIR.
Memelilerde Uçmanın Kökeni
Chiroptera Takımı içinde yer alan yarasalar, tüm memeliler içinde en özelleşmiş olanlarıdır ve tek uçan memelidirler. İki alt takıma bölünmüşlerdir. Megachiroptera, büyük, meyve yiyen yarasalardır ve bugün Eski Dünya tropikal bölgeleri ile Pasifik Bölgesi’nde bulunmaktadır. Microchiroptera, dünya çapında var olan küçük yarasalardır. Bu yarasaların çoğu geceleri böceklerle beslenirler. Tam karanlıkta bile onları algılayıp yakalayabilirler.
Şekil 13. Bir yarasa fosili iskeleti, Palaeochiropteryx.
Romer, Vertebrate Paleontology’den, University of Chicago Press’in izniyle.
Microchiroptera yarasaları bu yeteneğe sahiptirler çünkü meyve yiyen yarasalarda bulunmayan karmaşık bir ekolokasyon sistemiyle donatılmışlardır. Bu yarasalar nesnelerden yankılanacak tiz ses dalgaları yayarlar. Yankı yarasaya geri döner ve yarasanın çok hassas işitme organı tarafından alınır. Sonuç olarak beyin her saniye milyonlarca elektronik ileti göndermektedir ve beyin, bu elektronik mesajları bir görüntü haline dönüştürmektedir. Zifiri karanlık bir mağarada binlerce yarasa varken acaba bu yarasalar niçin diğer bir yarasadan gelen sesleri karıştırmazlar? Her yarasa, inanılmaz biçimde kendi sinyalini tanıma yeteneğine sahiptir ve görünüşe göre diğer tüm sinyaller yarasanın işitme sistemi tarafından filtre edilmektedir. Ne kadar ustaca yapılmış bir sistem! Microchiroptera yarasalarının bazıları balık ile beslenmek üzere tasarlanmışken, bazıları da sığır gibi büyükbaş hayvanların kanlarını emen “vampir” yarasalardır.
Burada yaratılış ile evrimi sınamak için güzel bir fırsata sahibiz ─ ileri derecede özelleşmiş bir memeli, uçamayan ve de muhtemelen böcekçil bir memeliden başlayarak sözde evrim yoluyla uçuş gücü kazanmıştır. Bu evrim süreci, milyonlarca yıllık bir zamanı kapsayacak ve kötü mutasyonlar okyanusundan tesadüfen ortaya çıkan çok sayıda, fazlasıyla nadir “iyi” mutasyon gerektirecektir. Bir parça değişime uğramış her “iyi” mutant, önceki olanın yerini alabilmek için, yüksek düzeyde yetenekli olan (eğer yüksek derecede yetenekli olmasaydı evrim geçirmezdi) bir rakiple, birçok nesil boyunca yarışmak zorunda kalacaktır. Her ara seviye sadece yaşamaya uygun değil, aynı zamanda kendinden önceki seviyeden daha üstün nitelikli olmalıdır. Bu süreç, karada yaşayan memeli atalarının ön üyelerinin, bir şekilde, her eldeki dört parmağın uzunlukları kademeli olarak artarken (başparmak temelde etkilenmeden), kanatlara dönüşmesini sağlamış olmalıdır. Başka bir dizi nadir “iyi” mutasyon ile kademeli olarak kanat zarları oluşmuş olmalıdır. Bu sürecin bir döneminde başka “iyi” mutasyonlar da adım adım uçuş kaslarını ve yarasaların özelleşmiş özelliklerinin gerektirdiği kirişlerin, sinirlerin ve kan damarlarının çok sayıdaki benzersiz düzenlenmesini geliştirmiş olmalıdır. Kafatası ve dişler uygun bir yolla değiştirilmiş olmalıdır ve microcheroptera yarasalarında, bir şekilde, rastgele genetik hatalar yoluyla, onların var oluş yapılarında bir seri hassas değişiklik gerçekleşmeli ve/veya yeni yapıların oluşumu, tam doğru bir sıra içerisinde gerçekleşen, başka bir seri çok nadir “iyi” genetik hata yoluyla başarılı bir sonuç vermelidir. Bu sayede bu yarasalardaki sonar ya da ekolokasyon sistemi ve aynı zamanda dikkatli bir şekilde, her yarasanın kendi yaydığı sinyali diğerlerinden ayırt edebilmesini sağlayan gerekli filtrasyon sistemi yaratılmalıdır.
Ş ekil 14.
ekil 14.
J. L. Jepsen tarafından yeniden yapılandırılmasının üzerine yerleştirilmiş olan en eski fosil yarasa fotoğrafı. 9 Aralık 1966’da yayınlanan Science’ın kapak sayfasından alınmıştır. 1966 telif hakkı, American Association for the Advancement of Science’a aittir.
Eğer evrim gerçekse, fosil kaydı bir dizi ara seviyenin en az bir kaçını göstermelidir. Örneğin, parmaklar uzarken ön üyelerin kademeli olarak kanatlara dönüşümündeki ve Microcheroptera yarasalarının ekolokasyon sistemini oluşturmak üzere kafatası ile işitme organının şekillenmesindeki ara seviyeleri göstermelidir. Fakat eğer yaratılış gerçekse, en eski yarasa fosili olduğuna inanılan canlı, %100 yarasa olmalıdır. Yarasaları, karada yaşayan memeli atalarına bağlayan hiçbir geçiş formu kanıtı bulunmayacaktır. Veriler hangi modele en iyi uymaktadır, yaratılış mı evrim mi? Verilere göre, yaratılış modelinin ezici üstünlüğü vardır. Aşağıda yer alan Carroll’a ait ifadeler her şeyi açıklamaktadır:
Yarasalar, günümüz memelilerinin en çok özelleşmiş olanları arasında yer alırlar. Tümü, becerikli uçuculardır ve böcekçil microcheroptera’lar, karanlıkta böcek avlamalarını mümkün kılan ileri düzeyde gelişmiş bir sonar sistemine sahiptirler. Yarasaların uçuş yapıları, pterosaur’larda da olduğu gibi, fosil kaydında ilk ortaya çıktıklarında zaten ileri düzeyde evrimleşmiş halde bulunuyordu. Erken Eosen katmanına ait en eski yarasa iskeleti olan Icaronycteris, neredeyse günümüz yarasalarından ayırt edilemez bir görünüşe sahiptir.59
Hiç kimse bunu daha iyi anlatamaz; ancak vurgulamak için devam edelim. Glenn Jepson’a göre, fosil kaydında yarasayla ilgili olan ve Icaronycteris’ten daha yaşlı herhangi bir canlı bulunamamıştır ve bu canlı temelde günümüz yarasalarıyla aynıdır.60
Son zamanlarda yapılan araştırmalar, Icaronycteris’in de günümüz microchiroptera yarasalarında bulunan sonar sistemine sahip olduğunu ortaya çıkarmıştır.61 Böylece yarasalar, fosil kaydında, hiçbir atasal ya da geçiş formu olmaksızın tümüyle gelişmiş biçimde ortaya çıkmaktadırlar ve bu canlılar, fosil kaydında sözde ilk ortaya çıktıkları zaman olan 50 milyon yıl öncesinden bu zamana kadar hiçbir değişime uğramaksızın aynı kalmışlardır. Elbette ki bu kanıt, evrim kuramıyla kesin bir zıtlık içinde bulunurken, yaratılış modelinin dayandırılmış olduğu tahminlerle tam bir uyum içerisinde bulunmaktadır. Pek çok yarasa fosiline sahip olduğumuz için evrimciler, yarasaların fosil kaydının yetersizliği gibi bir bahaneyi öne süremezler. Messel’de bulunan fosillerin çoğu yarasa fosilleriydi.62 Bugün, yarasalar, kemirgenler dışında, tüm memeliler arasında en doğurgan olanları temsil etmektedirler.
MEGACHIROPTERA YARASALARININ, İNSANLARA OLAN YAKINLIĞI, MICROCHEROPTERA YARASALARINA OLAN YAKINLIĞINDAN DAHA MI FAZLADIR? Avustralya Brisbane’deki Queensland Üniversitesi’nden John Pettigrew, megachiroptera’ların, yine yarasa olan microcheroptera’lardan daha çok primatlarla yakın akraba olduklarını ileri sürmüştür.63 Pettigrew’un bu fikrine katılan evrimciler, insanlar maymun (primat) oldukları için, meyve yiyen yarasaların, böcekçil yarasalar kadar ya da daha fazla, insanlarla yakın akraba olduklarına inanacaklardır. Pettigrew’a göre, büyük yarasaların karmaşık görme sistemleri, yani bir görüş etkisinin beyne iletilmesi biçimi, primatlarınkine benzer fakat küçük yarasalarınkinden farklıdır. Pettigrew, hem primatlarda hem de yarasalarda işlev gören sinirsel niteliklerin, bu iki farklı tür canlıda birbirinden bağımsız biçimde evrimleşebildiklerinden ciddi bir şekilde şüphelenmektedir.
Pettigrew’un görüşlerini eleştirenler, birbirinden farklı iki tane bağımsız evrimleşen canlının, hem büyük yarasalarda hem de küçük yarasalarda var olan aynı tür yarasa kanatlarına ulaşmasını hayal etmenin, bu iki tür yarasada bulunan görme sinir sisteminin bağımsız bir evrimsel kökene sahip olduğunu hayal etmekten daha zor olduğuna inanmaktadırlar.64 Yaratılışçı bilim adamları, sadece bir ya da iki geçiş formunun bulunmasının tartışmayı sona erdirmeye yeteceğini hatırlatmaktadır.
Kemirgenler, Yaratılış İçin Olumlu Kanıtlar Ortaya Koymaktadırlar
Rodentia Takımı, evrimciler için evrimsel çalışmalarda ideal hayvan grubu olması gereken bir takımdır. Kemirgen tür ve cinsleri sayı olarak, diğer tüm memeli takımlarının toplamından fazladır. Bu canlılar, hemen hemen tüm şartlar altında sağlıklı yaşayabilirler. Eğer bize geçiş formları sunabilecek bir canlı grubu varsa, bu grup elbette kemirgen grubu olmalıdır. Kemirgenlerin kökenleriyle ilgili olarak Romer şöyle demiştir:
Kemirgenlerin kökeni karanlıktır. Bu canlıların Paramys cinsinde, geç Paleosen’de ilk ortaya çıktığı sırada, takıma ait ayırt edici nitelikleri iyi gelişmiş, daha ilkel ama tipik gerçek bir kemirgenle karşı karşıya kalıyoruz. Bu canlılar tabi ki de bazı temel, böcekçil, plasentalı atalardan ortaya çıkmıştır ama hiçbir geçiş formu bilinmemektedir.65
Üstelik, temel kemirgen türleri arası geçiş formları fosil kaydında bulunmamaktadır. Örneğin Romer şöyle demektedir:
... kunduzlar tahminen ilkel sincapsı atalardan türemiştir fakat, bu tür formlar ve en yaşlı Oligosen kunduzumsuları arasında doğrudan bir ilişki kurabilecek hiçbir bağlayıcı tür bulunmamaktadır.66
Romer, Eski Dünya kirpileri olan Hystricidae hakkında konuşurken şöyle demektedir:
Miyosen’e kadar ve belki de Geç Oligosen’e kadar uzanan birkaç fosil formu bulunmaktadır; fakat bunlar, hystricid’ler ile başka kemirgen sınıfları arasında ne tür bir ilişki bulunduğuna dair hiçbir belirti göstermemektedir.67
“Kaya sıçanı” (Petromus) hakkında yorum yaparken Romer şöyle der: “Petromus’un ataları konusunda hemen hemen hiçbir şey bilinmemektedir.”68 Theridomyoidea üst familyası hakkında Romer şöyle demektedir: “Bugün, onların ataları ya da olası torunları hakkında hiçbir şey bilmiyoruz.”69 Romer, daha önceleri kemirgenler alt takımında, bugün ise farklı bir takımda, yani Lagomorpha’da yer alan “lagomorph’ların (tavşan çeşitleri), diğer plasentalı gruplara hiçbir yakın benzerlik göstermediğini ve en eski olarak bilinen formlarda bile takım nitelikleri çok iyi gelişmiş halde” 70 olduğunu kabul etmek zorunda kalmaktadır.
Böylece görüyoruz ki, evrim için, eğer evrim gerçekten de olmuşsa, mükemmel kanıtlar sağlaması gereken Rodentia takımı, yaratılış için güçlü kanıtlar ortaya koymaktadır.
Ünlü At Serisi
Kökenler sorusu göz önüne alındığı zaman atlar, en ilginç memeli gruplarından birini oluşturmaktadır. Hemen hemen tüm öğrenciler, ön ayaklarında dört parmağı bulunan, köpek büyüklüğünde bir “at” olan Hyracotherium (Eohippus)’la başlayan, düz bir evrim çizgisi boyunca üç ayaklı türlere evrimleşen ve modern tek parmaklı Equus ile son bulan at “evrimi” hikayesini duymuştur. Birdsell, genelde atların evrimini kabul ederken şunu da ilan eder: “Bu hikayenin büyük kısmı doğru değildir...”71 Başkaları da aynı görüşü paylaşmaktadır. Örneğin George Gaylord Simpson, birkaç öğrenci neslinin, at evriminin gerçek anlamı hakkında yanlış bilgilendirildiğini ifade etmiştir.72 Bu yazarlar at evriminin, genellikle betimlenenden çok daha karmaşık olduğuna, belki de bir ağaçtan çok bir dizi çalıya benzediğine inanmaktadırlar.
Bizim açımızdan atların familya ağacı sadece, eşit olmayan parçaların bir araya getirildiği bir çeşit senaryo gibi görünmektedir. Örneğin, at olmayan ve her ayağında beş toynak bulunan bir atanın (Condylarth’a ait olduğu varsayılan atanın), geride üç, önde dört ayak parmağı bulunan Hyracotherium’a dönüşümünü belgeleyen hiçbir ara form bulunmamaktadır. Ne dört parmaklı Hyracotherium ile üç parmaklı Miohippus arasında ne de dallardan otlanmaya yarayan dişlerle donatılmış Miohippus ile üç parmaklı ve yüksek taçlı öğütücü dişlerle donatılmış Merychippus arasında hiçbir geçiş formu bulunmamaktadır. Son olarak, Equus gibi tek toynaklı otlayanların, üç toynaklı otlayanlardan kademeli evrimsel olarak dönüştüğünü gösteren hiçbir ara seviye olmaksızın aniden ortaya çıkmaktadırlar.
Aynı şekilde Birdsell, bu hikayeyi bize aşağıdaki gibi anlatmaktadır. Bir evrimcinin dönüşüm hakkında “birdenbire,” “aniden,” “hızlı” kelimelerini kullanarak, hiçbir geçiş formu bulunamadığını ima edişine dikkat edin:
Ayak yapısının evrimi, kademeli değişimden ziyade, hızlı ve aniden oluşan değişimlerle ilerlemektedir. Eohippus minyatüründe gösterilen ayak biçiminin, daha büyük ve hep üç parmaklı olan Miohippus’un ayağına dönüşümü öylesine ani olmuştur ki fosil kalıntılarında tek bir iz bile bırakmamıştır... onların ayak yapıları, tabanın kaybolduğu ve iki taraftaki parmakların artık işlevsiz hale geldiği üç parmaklı ayak yapısına çok hızlı dönüşmüştür. Sonunda Pliyosen’de, modern tek parmaklı otlayıcılara doğru yönelen çizgi, her bir ayağın her iki yanındaki parmakların hızla kaybına doğru yönelmiştir.73
Daha sonra Birdsell bu evrimin kademeli değil de hızlı sıçramalar yoluyla ilerlediğini söyleyerek sözlerine devam etmiştir.
1980 Kasım’ında 150 evrimci, kademeli evrim kuramını ya da yeni Darvinci evrim kuramını eleştirmek ya da savunmak için, Chicago’daki Field Museum of Natural History’de dört gün boyunca toplandılar. Yeni Darvinci evrim mekanizmasını eleştirenler, fosil kaydında kademeli değişimi gösteren ya hiç ya da çok az kanıt bulunduğunu iddia etmişlerdir. Doğal olarak, ileri sürülen at evrimi hikayesi de tartışıldı. Bu toplantı hakkında yazdığı raporunda Boyce Rensberger şöyle demiştir:
Halk arasında anlatılan at evrimi örneğinin, yani bundan 50 milyon yıl önce yaşayan tilki büyüklüğündeki dört toynaklı canlıların, kademeli bir değişim dizisiyle bugünkü çok daha büyük, tek toynaklı atlara dönüşümünü ileri süren at evrimi örneğinin yanlış olduğu uzun zamandan beri bilinmektedir. Kademeli bir değişim göstermeksizin her ara tür fosili, tümüyle birbirinden farklı olarak ortaya çıkmış, değişmeden kalmış ve sonra da yok olmuştur. Geçiş formları bilinmemektedir.74
Yukarıda anlatılanlardan çıkarılacak sonuç gayet açıktır: Evrim kuramının gerektirdiği süreklilik, fosil kaydı tarafından belgelenememektedir. Diğer yandan Rensberger’in de söylediği gibi, at türlerinin her biri tümüyle gelişmiş biçimde ortaya çıkarlar ve fosil kaydında yok oluncaya kadar hiçbir değişim göstermezler. Bu, yaratılış modelinde öngörülen kanıt çeşitlerindendir.
Kuzey ve Güney Amerika toynaklılarını karşılaştırdığımızda oldukça açık ve şaşırtıcı bir gerçek görebiliriz. Şekil 15’te gösterilen diziyi hepimiz daha önceden görmüşüzdür. Bunlar, (a) “Eohippus”un (b) yan toynakları kısalmış Merychippus’un ve (c) modern Equus’un arka ayaklarıdır.
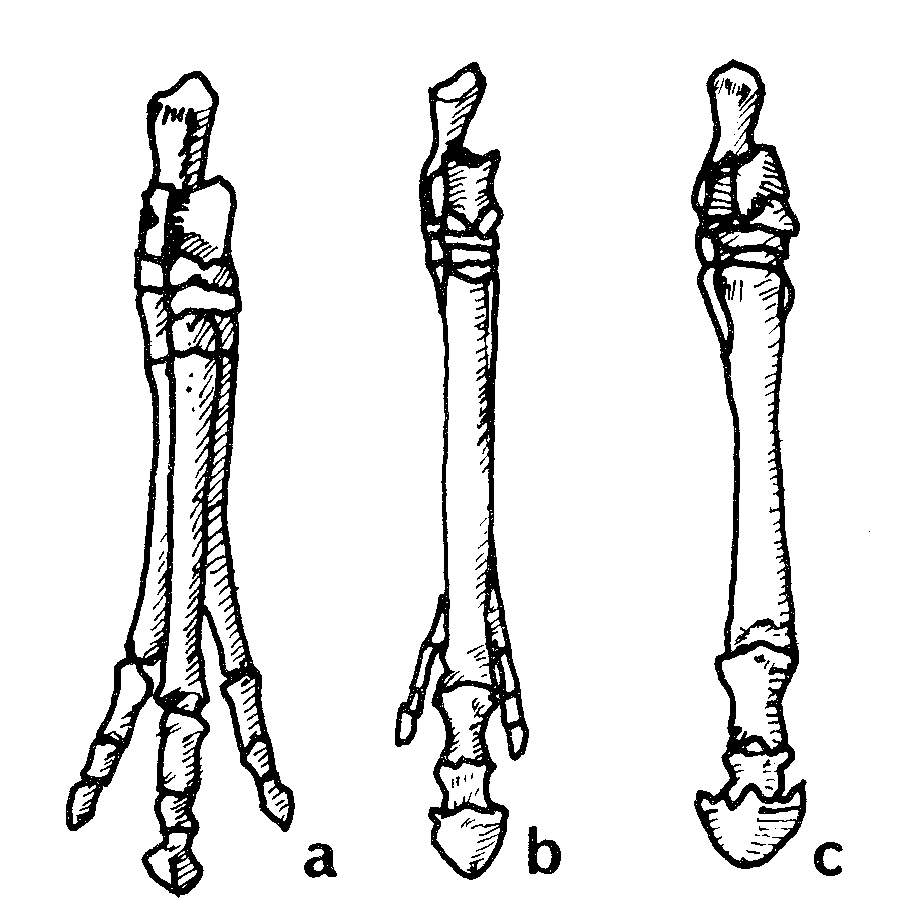
Şekil 15. a) “Eohippus”; b) Merychippus; c) Equus’un arka ayakları (pes’leri).
Şimdi Şekil 16’ya bakalım. Şekilde gösterilenler, Güney Amerika toynaklılarının (Litopterna Takımı) arka ayaklarıdır, (a) Macrauchenia; (b) Diadiaphorus; ve (c) Thoatherium. Şekilde, üç toynaklı olan Macrauchenia, yan toynakları kısalmış üç toynaklı olan Diadiaphorus ve tek toynaklı olan Thoatherium görülmektedir. Romer, Thoatherium’un, kamış kemikleri modern atlarınkinden daha kısa olan tek toynaklı gerçek bir attan çok, at benzeri bir canlıya benzediğini söylemektedir.75
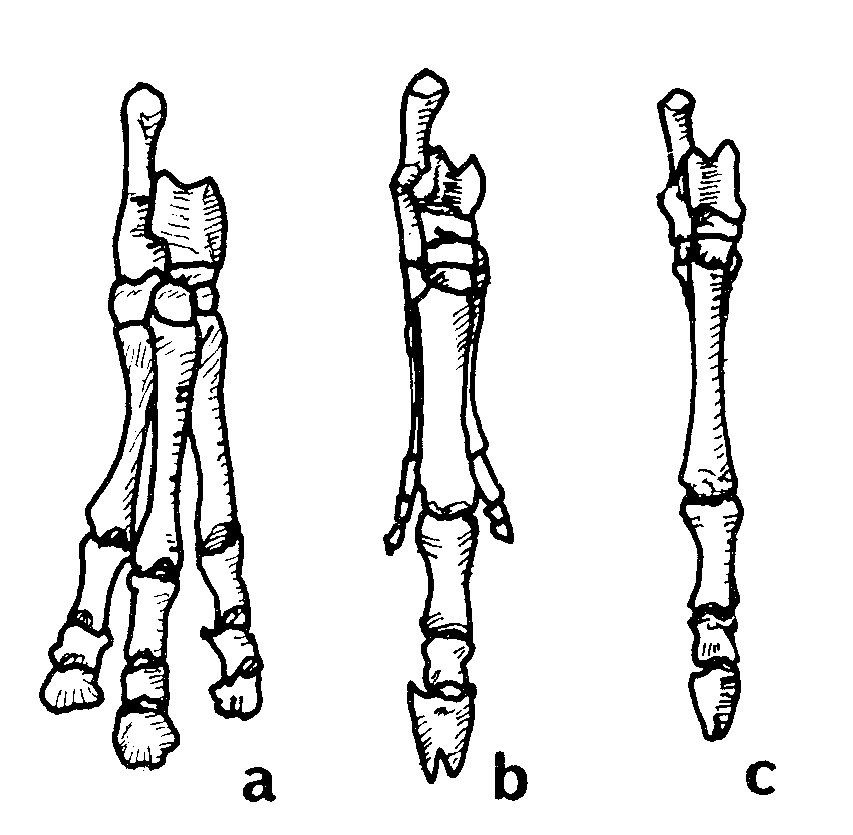
Şekil 16. LitopternaTakımı’ndan Güney Amerika toynaklılarının arka ayakları, a) Macrauchenia, b) Diadiaphorus, c) Thoatherium.
Bunlar da güzel ve mantıklı başka bir evrim serisi oluşturmuyor mu? Hayır, çünkü zaten onlar bu sırayla fosil katmanlarında var olmadılar! Yan toynakları kısalmış üç toynaklı olan Diadiaphorus ve tek toynaklı olan Thoatherium'un her ikisi de Miyosen Çağı’nda yaşamıştır. Üç tam arka ayak toynağına sahip olan Macrauchenia ise, jeolojik sütuna göre Miyosen’den sonra gelen Pliyosen Çağı’na kadar bulunmamıştır ve bu canlının fosilleri Pleistosen kayaları arasında da bulunmaktadır. Gerçekte tek toynaklı Thoatherium’un, üç toynaklı Macrauchenia Pliyosen’de ortaya çıkmadan önce, Miyosen’de yok olduğu söylenmektedir.
Eğer evrimciler fosil kanıtlarının ve jeolojik zamanlarla ilgili varsayımlarının onlara yol göstermesine izin verirlerse, Güney Amerika’daki tek toynaklıdan, yan toynağı kısa üç toynaklının çıktığına ve bu canlıdan da üç tam toynağa sahip bir toynaklı çıktığına inanacaklardır. Bu, Kuzey Amerika atlarında gerçekleştiği varsayılan olaylar dizisiyle tam bir zıtlık oluşturmaktadır. Böyle evrimsel olaylar dizisi öneren hiçbir evrimci tanımıyoruz, peki neden? Belki de Kuzey Amerika atlarındaki bu üç toynaklılıktan tek toynaklılığa geçiş, evrim çevrelerinde öylesine herkes tarafından kabul edilir hale geldi ki, hiç kimse tersine dönüşü önermeye cesaret bile edemiyor. Ve tabi ki Güney Amerika’da da, Kuzey Amerika’da bulunandan daha fazla geçiş formu kanıtı bulunmamaktadır.
Oregon’un kuzeydoğusundaki John Day ilçesinde, Rattlesnake Oluşumunda üç toynaklı Neohipparion atı ile birlikte tek toynaklı Pliohippus atı bulunmuştur.76 Bu ikisinin arasında hiçbir geçiş formu bulunamamıştır. Ayrıca, başka yerlerde Merychippus gibi “ilkel” cins türleri, “gelişmiş” türleri içinde barındıran oluşumlardan sözde daha genç jeolojik oluşumlarda bulunmaktadır.77
Gerçekte, evrimciler tarafından yaygın olarak ileri sürülen üç toynaklı atların tek toynaklı atlara evrimleştiği fikri, kanıtlar tarafından desteklenmemektedir. Bugün yalnızca tek toynaklı atların yaşadığı doğrudur fakat bunun nedeni, bu atların üç toynaklı atlardan evrimleşmeleri ve evrimsel anlamda daha üstün nitelikli oldukları için onların yerini almaları değildir. Üç toynaklı atlar ve tek toynaklı atlar, Kuzey Amerika’da, sık görüldüğü gibi, aynı zamanda ve birlikte yaşamışlardır. Örneğin Nebraska’nın kuzeydoğusunda, kuş, sürüngen ve memeli çeşitlerinin bulunduğu dikkate değer bir fosil mezarlığı vardır. Küçük bir gölü temsil ettiği söylenen bu tortu, Nebraska Eyalet Müzesi’nin omurgalı paleontoloji müdürü olan Michael Voorhies tarafından keşfedilmiştir.78 Buradan, neredeyse tüm halde, 200’den fazla fosil iskeleti çıkarılmıştır. Voorhies, bu canlıların, ani bir afetten çok, aylarca devam eden volkanik hareketler sonucu kül yağmasıyla öldüklerine inanmaktadır. Bu zengin memeli faunasında, beş at türü bulunmuştur: Pseudhipparion gratum, Cormohipparion occidentale, Protohippus supremus, Astrohippus’un Neohipparion türü ve Dinohippus.79 Dinohippus çeşitleri, üç toynaklı ve tek toynaklı atlardan oluşmaktaydı. Bu yüzden MacFadden şöyle demektedir:
Nebraska’nın kuzeydoğusundaki Ashfall (Kül yağmuru) Fosil Yatakları’nda, mükemmel derecede korunmuş ilkel Dinohippus Voorhies’in Pliohippus’u (1981) popülasyonunun son zamanlardaki keşfi,… bazı bireyler üç toynaklıyken, bazılarınınsa tek toynaklı olduğuna işaret etmektedir (Voorhies (1981)). Üstelik bu durum, önceden sadece tek toynaklı oldukları düşünülen başka ilkel at türleri için de geçerli olabilir…80
Daha sonra MacFadden bize şöyle demektedir:
Böylece uyum sağlamış en az üç farklı at fosili grubu, Orta ve Geç Miyosen boyunca çeşitli postcranial şekillerle bir arada yaşamışlardır. Anchitere’ler, ayak tabanıyla birlikte ilkel üç toynaklı ayak yapılarını korudular. Gelişmiş üç toynaklı atlar, pek çok yönden tek toynaklı formlara benzeyen toynaklı seviyesinde ayaklar evrimleştirdiler. Sonunda, bu zaman boyunca, daha sonra Equus’ta oluşan hareket sistemlerine sahip tek toynaklı atlar ortaya çıktı.81
Tek bir zaman içerisinde, bir arada yaşamış olan ve “ilkel” üç toynaklı atlar, “gelişmiş” üç toynaklı atlar ve tek toynaklı atlar olmak üzere üç farklı at grubu vardı. Evrimcilerin yayınlarında, bu atları, sıraya konmuş bir halde görürüz ve bu yayınlar, “ilkel” üç toynaklı atların, “gelişmiş” üç toynaklı atlara ve “gelişmiş” üç toynaklı atların da sonunda tek toynaklı atlara evrimleştiğini ima etmektedirler. Açıkça bu böyle olmamıştır. Daha önceden de belgelendiği gibi, üç toynaklı bir atın, tek toynaklı bir ata evrimleştiğini gösteren hiçbir geçiş formu bulunmamaktadır. Gerçekte bu canlılar, bir arada yaşıyorlardı. Güney Amerika’daki litoptern’ler hakkında yazılmış olan kısa bir bölümde MacFadden şöyle demektedir:
… onlar, Güney Amerika’da soyu tükenmiş olan bir otçul grubuydu; Kuzey Amerika’daki atlardan yaklaşık 8 milyon yıl önce Erken Miyosen sırasında tek toynaklı oldular.82
MacFadden, üç toynaklı litoptern’lerin, sözde tek toynaklı çeşidin nesli tükendikten sonra ortaya çıkması hakkında tek bir söz bile söylemekten kaçınmıştır.
Walter R. Barnhart, genellikle at olarak adlandırılan bu canlıların (Equidae) kökeni ve çeşitliliği ile ilgili bilimsel yayınlar üzerinde yaptığı yoğun çalışmalardan sonra, birçok şaşırtıcı sonuca ulaştı.83 Araştırmaları onu, karışık cinslerden meydana gelen fauna gruplarının görünmesinin, sadece göreli küçük değişimler ya da sık sık söylendiği gibi “mikro evrim” geçirme niteliği taşıyan, birlikte çiftleşen popülasyonların var olduğunu ima ettiği sonucuna ulaştırmıştır. Barnhart, taksonların jeolojik mesafesinin, tüm at cinslerinin Miyosen’de bulunduğunu gösterdiğini ve evrimin gerektirdiği jeolojik ilerlemenin asla var olamadığını duyurmuştur. Barnhart’ın çıkardığı sonuç, equini fosil grubunun yalnızca üç cinsten oluştuğuydu: Hyracotherium, Mesohippus ve Equus.
Barnhart, Hyracotherium’un altına Orohippus ve Epihippus’u da tür olarak dahil etmektedir. Miohippus, Anchitherium, Hypohippus, Megahippus, Archaeohippus ve Parahippus’un, Mesohippus’a ait çeşit ya da türler olarak; Merychippus, Hipparion, Stylohipparion, Neohipparion, Nannippus, Calippus, Pliohippus, Hippidion, Orohippidium ve Parahipparion’un hepsinin Equus altındaki türler olarak gruplanması gerektiğini iddia etmektedir. Ayrıca Barnhart, evrim kuramının gerektirdiği geçiş formlarının yokluğunu da ifade etmektedir. Fosil atların (Equidae) makro evrimi gösteren bir evrim serisi oluşturduğu fikrinin, kanıtların yakın ve dikkatli incelenmesiyle yalanlandığını iddia etmektedir.
Hyracotherium (Eohippus) gerçek bir at mıydı? Hyracotherium, Kuzey Amerika’da “Eohippus” çıkarılmadan önce Avrupa’da keşfedilmiştir ve Hyracotherium cins adı, aynı zamanda bu canlıyı keşfeden ünlü bir İngiliz anatomist ve paleontolog Richard Owen tarafından verilmiştir. Daha sonra Kuzey Amerika’da başka örnekler keşfedildi ve bu örneklere Eohippus cins adı verildi. Daha sonraları, Kuzey Amerika’da bulunan örneklerin, Hyracotherium ile aynı cinsten oldukları kararına varıldı. Hyracotherium adı daha önce verildiği için öncülüğü taşımaktadır ve Eohippus, bu canlılar için geçerli bir isim değildir. Ancak, Eohippus adının yaygın kullanılmasının nedeni, şüphesiz, Eohippus isminin “ilkel at” anlamına gelmesi ve Owen’in bu canlının Hyrax (kaya tavşanı, kır sıçanımsı) cinsi canlılara olan benzerliği nedeniyle ona Hyracotherium adını vermesidir.
Hyracotherium ya da “Eohippus”, ne habitat ne de morfolojik açıdan günümüz atlarına benzememesine rağmen, atların kökenini oluşturmak üzere Amerikalı paleontolog Marsh ve diğerleri tarafından seçildi ve canlının bu konumu, Thomas H. Huxley tarafından New York şehrinde verilen bir konferanstan ve Marsh’ın çalışmalarının yayınlanmasından sonra, hem halk arasında hem de bilimsel statüde yer edindi.84
Nilsson, Hyracotherium’un, atlara çok az ya da hiç benzememesiyle birlikte, habitat ve morfolojik açıdan yaşayan Kaya tavşanı cinsine ait canlılara ne kadar benzer göründüğünü işaret etmektedir.85 Kaya tavşanı (hyrax) da Hyracotherium gibi ön ayağında dört, arka ayağında üç toynağa sahiptir. Bu iki canlının yan dişeri, pek çok benzerliği paylaşmaktadır ve bu dişler, atlarınkinden çok, gergedanlarınkine benzemektedir. Kaya tavşanının habitat ve yaşam tarzı da Hyracotherium’da varsayılana benzemektedir. Nilsson, Hyracotherium’un hiçbir bakımdan bugünkü atlara benzememesiyle birlikte görünürde, dikkate değer derecede bugünkü kaya tavşanına benzemekte olduğunu iddia etmektedir.
Başka insanlar daHyracotherium’un atlarla olan akrabalığından şüphelenmektedir. Örneğin Kerkut şöyle der:
İlk olarak, Hyracotherium’un atların atası olup olmadığı kesin değildir. Mesela Simpson (1945) şöyle der: “Matthew, Hyracotherium’un (Eohippus da dahil), tapirimsi, gergedanımsı...vs.den çok daha fazla ata benzemeyecek kadar ilkel olduğunu göstermiş ve ısrar etmiştir. Fakat onu at grubunun kökenine yerleştirmek artık yaygınlaşmıştır.”86
Diğer bir deyişle Hyracotherium’unata olan benzerliği, onun tapire ya da gergedana olan benzerliğinden fazla değildir ve onun için, bu canlı aynı şekilde gergedanın ya da tapirin atası olarak da seçilebilirdi. Öyleyse, atların soy ağacını oluşturan insanların tarafsızlığının başlangıçtan itibaren sorgulanabilir olduğu ve tüm at familyasının üzerinde durduğu ağacın kökü olan “at”ın da, asla bir at olmadığı görülmektedir.
Üstelik, George Gaylord Simpson, dünyanın hiçbir yerinde, evrimcilerin Hyracotherium’un ve böylece tüm atların atası olarak önerdiği memeli takımı olan Condylarthra ile Hyracotherium arasındaki büyük boşluğu doldurabilecek hiçbir fosil izi bulunmadığını itiraf etmiştir. William J. Morris, Hyracotherium’dan Equus’a kadar olan atların soy oluşu konusundaki inançlarını dile getirirken şöyle demiştir:
… Hyracotherium’un en erken popülasyonları ile olası condylarth atalar arasında soy oluşsal bir halka bulunamamıştır. Ve dahası, Hyracotherium’un Eosen Çağında aniden, hatta Geç Paleosen hayvanlarının çok iyi bilinen bölgelerinde bile ortaya çıkışı, rahatsız edici bir durumdur.87
Atlar konusunda Kerkut’un kitabından bu yana onun kararını önemli derecede değiştirebilecek kapsamlı hiçbir çalışma yayınlanmamıştır:
Bazı yönlerden bu, Osborn (1937, 1943) tarafından önerilen Hortumlular(Proboscidea) takımının evrim modeli kadar karmakarışık görünmektedir; çünkü Hortumlular için “hemen hemen bilinen hiçbir form diğerinin bir soyu olarak düşünülememektedir; her alt gruplamanın birbirinden tümüyle ayrı ve genellikle Erken Eosen ya da Geç Tebeşir’deki kuramsal ortak atalardan gelen hiçbir bilinen ara seviye bulunmaksızın çıktığı varsayılmaktadır” (Romer, 1949).88
Eğer gerçekten de “atların evrimi” böylesine karmakarışık ve yarım yamalaksa, bu klasik evrim savının gerçek bir geçerliliği yoktur. Diğer yandan gerçek kanıtlar yaratılış modeli ile tam tamına örtüşmektedir.
Deniz Memelileri
Evrimciler, evrim kuramlarını doğrulamak için geçiş ya da ara seviye formlarını bulmak için çırpınmaktadırlar. Bu ara seviyelerinin olmaması, balinaların, yunus balıklarının ve diğer deniz memelilerinin kökenleriyle ilgili konularda da çarpıcı biçimde doğruluk taşıyan bir durumdur. Romer, yarı toynaklılar (kaya tavşanları, filler, deniz ayıları) hakkındaki toparlayıcı sözlerinden birinde şöyle demektedir: “Kaya tavşanları, hortumlu memeliler ve Sirenia takımına ait memeliler fosil kaydında ilk ortaya çıktıkları zamanda zaten farklı gruplar halindeydiler.”89 Olson, eğer deniz sürüngenlerinin atalarını ararsak, deniz–kara arası geçiş formlarıyla ilgili hiçbir şey aklımıza gelmez demektedir.90 Bu sözler, fok balıklarını, yunus balıklarını ve balinaları kapsamaktadır.
Balinalar hakkında konuşan Colbert şöyle demektedir:
Bu memeliler, eski bir kökene sahip olmalıdırlar; çünkü, fosil kaydında, balinalar ile Tebeşir’e ait atasal plasentalılar arası hiçbir ara seviye formu görünmemektedir. Yarasalar gibi balina ve balina benzerleri Erken Üçüncü Zamanda (Tersiyer), oldukça özelleşmiş bir yaşam biçimi için gerekli olan, temel memeli yapılarının geçirdiği etkileyici modifikasyonlara tümüyle uyum sağlamış biçimde ortaya çıkmıştır. Gerçekte balinaların memeliler içindeki konumu yarasaların konumundan çok daha ayrıdır; tümüyle yalnızdır.91
Wursig yunus balıklarının, bugünkü sığırlar, domuzlar ve bufalolar gibi çift sayıda toynaklılara benzer kara memelilerinden evrimleşmiş olabileceklerini ileri sürmüştür.92 Ara seviye formlarının neye benzediklerini, inek, domuz ya da bufalolarla başlayarak gözümüzde canlandırmamız oldukça eğlencelidir.
Eskiden National Geographic Dergisinde yer alan balinalar hakkındaki makale ile birlikte resimli bir ekte bize şöyle denmiştir:
Görünüşe göre balinaların üstün büyüklüğü, altmış milyon yıl önce, yiyecek ya da sığınak arayışında olan kıllı, dört bacaklı memelilerin suya girme cesaretini gösterdikleri zaman başlamıştır. Değişimler, çağlar boyunca yavaş yavaş meydana gelmiştir: arka bacaklar kaybolmuş, ön bacaklar yüzmede kullanılan paletlere dönüşmüş, kıllar balinanın kalın, yumuşak yağ tabakasını oluşturmuş, burun delikleri başın üst kısmıyla yer değiştirmiş, kuyruk yassılaşarak palete dönüşmüş ve kaldırma gücü olan su ortamında vücut dev bir büyüklüğe ulaşmıştır.93
Geçtiğimiz yıllarda evrimcilerin, kıllı, dört ayaklı hayvanların gerçekten suya girme cesaretini gösterdiklerinin ve milyonlarca yıl süren kademeli bir süreç sonucu balinalara evrimleştiklerinin gerçek kanıtları olarak değerlendirdikleri fosilleşmiş canlılarla ilgili olarak bir dizi keşif yapılmıştır. En çok sevdiği uğraş yaratılışçılara saldırmak olan Stephen Gould, bu bulguları övgü dolu sözlerle açıkladığı bir makaleyi hemen yayınlamıştır.94 Gould evrimcilerin, fosil kaydının, evrim kuramının gerektirdiği geçiş formlarını sağlamada başarısızlığa uğramış olduğunu iddia eden yaratılış yanlıları karşısında nihayet bir zafer kazanmış olduklarına inanmakta ve şöyle demektedir:
Bilimin halka açıklanması için daha iyi bir örnek ya da direnç gösteren yaratılışçılara karşı daha doyurucu ve akılcı temellere dayandırılmış bir siyasi zafer öyküsü düşünemiyorum.
Makalenin önceki kısımlarında Gould şöyle demişti:
Sözlü hilekarlıklarla beyazı siyah, siyahı beyaz yapmayı bilen bu bağnazlar, hiçbir konuda asla ikna olmayacaklardır fakat Ambulocetus, onların kendi varsayımlarına göre imkansız diye duyurdukları canlının ta kendisidir.
Yaratılışçı bilim adamları, aslında 120 trilyon bağlantı ve 12 milyar beyin hücresinden oluşan insan beyninin asıl kökeninin eninde sonunda yalnızca, hidrojen ve helyum gazlarının karışımı ile yer çekimsel enerjinin toplamı olduğuna inanan evrimcilerin, evrim kuramları gerekli gördüğünde siyahın beyaz, beyazın siyah olduğuna inanabilen kişiler olduklarını hatırlatmaktadır. Alice Harikalar Diyarındaki Kupa Kızı’nın da söylediği gibi, eğer bir kişi yeteri kadar alıştırma yaparsa, kahvaltı etmeden önce altı tane imkansız şeye rahatlıkla inanabilir.
Bugün evrimciler arasındaki ortak karar, tüm deniz memelilerinin olası atasının kıllı, dört ayaklı etçil memeli Mesonyx olduğudur. Bu görüş, en çok dişlerle ve kafatası biçimi ile ilgili kanıtlara dayandırılmıştır. Mesonychid’ler, herkesin bildiği kadarıyla su içme ihtiyacı dışında asla suya yaklaşmayan kurt benzeri, toynaklı etçillerdi. Carroll şöyle demektedir: “Mesonyx, bir kurt büyüklüğüne ve vücut ölçülerine sahip ve belki de kurt gibi yaşayan bir canlıydı.”95 Şimdi bu kurdu ya da kurt benzeri canlıyı okyanusun içinde bırakalım ve onun evrim senaryosuna göre bizleri ne tarafa yönlendireceğine bir bakalım.
İlk haber, yeni bir Basilosaurus fosilinin keşfi ile ilgiliydi. Basilosaurus, 19. yy’ın başlarında, Eosen kayaları arasında bulunan, yılan benzeri büyük bir omurgalıydı. “Kral kertenkele” anlamına gelen ismi ona, 1834 yılında R. Harlan tarafından verilmişti. Harlan’ın ona bu ismi vermesinin nedeni, onun bir sürüngen olduğu düşüncesinde olmasıdır. Daha sonraları başkaları onun bir memeli olduğunu düşünmüş fakat bu canlının ismi aynı kalmıştır. 1990’da Gingerich, Smith ve Simons, 1987 ve 1989’da, Mısır’ın orta kuzeyindeki çölündeki Reuglodon Vadisi’nde 243 kısmiBasilosaurus iskeletini konumlandırdıklarını duyurmuşlardır.96 Bu canlı, biraz büyük ön ayak ve bacaklara sahip olmasının yanı sıra böyle bir canlıya göre çok küçük olan eksiksiz bir çift arka bacağa da sahipti. Bu kişiler, çoğu eklemin iyi geliştiğini, diz kapağı ve topuk tümseğinin güçlü kasların eklenebileceği kadar büyük olduğunu ve dizlerin karmaşık bir kilitleme mekanizmasına sahip olduğunu söylemelerine rağmen, arka üyelerin yüzmeye yardımcı olamayacak kadar küçük olduklarına ve karada vücudu destekleyemediğine inanmaktadırlar. Bu yüzden, arka üyelerin büyük olasılıkla çiftleşmeye yardımcılar olarak kullanıldıklarını ortaya atmışlardır. Evrimciler, günümüz balinalarının birkaçında bulunan leğen kemiklerinin, karasal atalardan kalmış ve körelmiş bir yapı olduğunu varsaymaktadırlar. Ancak, bu leğen kemikleri, kullanılmamak üzere körelmiş yapılar değillerdir. Bunlar, çok önemli bir işlevi yerine getirmektedir. Günümüz balinalarının sahip oldukları leğen kemiği, üreme organlarının sıkıca tutulması görevini üstlenmektedir.97
Basilosaurus bir balina ya da balina olma yolunda bir canlı mıydı? Balinayı balina yapan şey nedir? Bu tümüyle insanların kendi yorumlarında kullandıkları varsayımlara bağlıdır. Bugün, ön ve arka üyelere sahip hiçbir balina yoktur. Acaba bir balina güçlü ön ve arka bacaklarla ne yapıyordu? Bu canlı her ne olursa olsun, yaratılışçı bilim adamları Basilosaurus’un, kurt ya da kurt benzeri bir hayvanla bir balina arası bir canlı olduğuna kesinlikle inanmamaktadırlar.
1983’te Gingerich ve meslektaşlarının yayınladıkları98 makaleye dayanan tüm dünya gazetelerinin manşetleri, balinalar ile onların kuramsal kara memelisi atası olan toynaklı memeli etçil Mesonyx arasında bir halka oluşturan ve ilkel olduğu söylenen bir balinanın keşfini haykırıyordu. Bu fosil kalıntıları sadece, kafatasının arka kısmı, iki altçene parçası ve birbirleriyle bağlantısı olmayan alt ve üst yan dişlerden meydana geliyordu. Bu canlı, Pakicetus inachus olarak adlandırıldı.
Bu fosil kalıntıları, kırmızı nehir tortuları arasında, yani akarsular tarafından üretilen ve demir madenlerinden süzülen materyal ile renklenen çökelimler arasında bulunmuştur. Böylece bu oluşum, karasal ya da kıtasal bir çökelimdir. Pakicetus’la ilgili fosil kalıntılarının çoğu kara memelilerine aittir. Memelilere ait olmayan kalıntılar, salyangoz, balık (özellikle yayın balığı), kaplumbağa ve timsah gibi başka karasal kalıntılar içermektedir. Bu kanıtlar, balina ya da balina benzeri bir canlı için ümit edilen bir deniz ortamından çok nehirsel ve kıtasal bir ortamı göstermektedir. Pakicetus’un, ne su altında gelen sesin yönünü ayırt ederek duyabildiğine, ne de dalma esnasında basıncı korumak için orta kulakta damarlar çıkmasına dair hiçbir kanıt bulunmadığı için, duyu mekanizmasının bir balinanınkinden çok, bir kara memelisine benzemesi oldukça dikkate değerdir. Yazarlar dişlerin, büyük olasılıkla sert sebzelerle, yumuşakçalarla ve leşlerle karınlarını doyuran mesonychid’lerinkine benzediğini söylemişlerdir. Bu kanıta göre, deniz memelileriyle hiçbir ilişkisi bulunmayan Pakicetus’un kara memelisinden başka bir şey olmadığı anlaşılmaktadır.
Daha yakın bir zamanda Ocak 1994’te Science’de yayınlanan bir makalede, kara memelileri ile deniz memelileri arası bir halkanın keşfinin mümkün olabileceği konusunda bir iddia yer aldı.99 Bu makale, bir kez daha, Amerika’da yayınlanan gazete başlıklarının temelini oluşturdu. Örneğin Cleveland şehrinde 16 Ocak 1994 tarihinde yayınlanan Plain Dealer adlı gazete, büyük bir iddia ile “Fosilin Yürüyen Bir Balinaya Ait Olduğu Düşünülüyor – Bu Canlı Kayıp Bir Halka Olabilir” başlıklı bir makaleye yer vermiştir. Balinalar karada yürümedikleri için, şüpheci kimseler derhal bu canlı ne olursa olsun onun bir balina olarak nitelendirilmesinin esasını sorgulamaya başlayacaklardır. Gerçekte, Science’in aynı baskısında asıl bilimsel rapor olarak yayınlanan açıklamada yazar şöyle demektedir: “Yazarlar, eski balinaların karada yürüme yetenekleri olan canlılardır şeklindeki, görünüşte mantık dışı olan yargılarına bazı kanıtlar sağlamaktadırlar.”100 Araştırmacılar bulgularına ambulare (yürümek), cetus (balina) ve natans (yüzen) kelimelerinin bir arada toplandığı Ambulocetus natans ismini verdiler. Böylece onlar bu canlının hem karada yürüdüğüne hem de suda yüzdüğüne inanmaktadırlar. Yazarlar raporlarında şöyle dediler: “Ambulocetus, günümüz memeli deniz hayvanlarından farklı olarak, büyük ihtimalle günümüz büyük fok balıklarınkine ya da kürklü fok balıklarınkine benzer bir şekilde kesinlikle karada yürüme yeteneğine sahiptiler. Bu canlı, suda, günümüz fok balıklarının, su samurlarının ve diğer memeli deniz canlılarının hareket yönlerini birleştirmişti ... Bu şekilde Ambulocetus, deniz memelileri ve kara memelileri arasında hayati bir ara seviye teşkil etmektedir.” 101
Kuzeydoğu Ohio Tıp Fakültesi’nde anatomi dalında yardımcı profesör olan Hans Thewissen, Harvard Üniversitesi’nde anatomi profesörü olan Tasseer Hussain ve Pakistan Jeolojik Araştırma Kurumu’nda jeolog olarak görev yapan M. Arif’in bu fosili, Pakistan’ın İslamabat şehrinin batısında, tepelerde yapılan 1992 kazıları sırasında tesadüfen buldukları bildirilmiştir. Plain Dealer adlı gazetede, yayınlanan makalenin yanı sıra bu fosilin iyi bir resmi de bulunuyordu. Yaratılış Araştırma Enstitüsü (Institute for Creation Research) çalışanlarından bazıları bu resme, Thewissen ve çalışma arkadaşlarının bu canlıyı bir balina olarak isimlendirdiklerini bilerek baktıklarında, doğal olarak, uçan bir fil ya da yürüyen bir balina görmedikleri için bu resmin balina olmasına şüpheyle yaklaşıyorlardı. Thewissen ve meslektaşları yazdıkları makalede Ambulocetus’un yaklaşık 300 kg. ağırlığında, bir erkek büyük fok balığıyla hemen hemen aynı boyutlarda olduğunu ve güçlü bir radyus ile ulna’ya (üst önkolda bulunan iki kemik) sahip olduğunu söylemişlerdir. Önkol yapısının, üç başlı kaslar (triceps) yoluyla güçlü bir biçimde uzamasına izin veren bir yapıda olduğunu ve günümüz deniz memelilerinden farklı olarak dirsek, bilek ve parmak eklemlerinin de yağlanmış ve esnek bir yapıda olduğunu bildirmişlerdir. Bu canlıların beş parmaklı elleri uzun ve genişti. Uyluk kemiği, kısa ve kalındı ve ayakları kocamandı. Ayak parmaklarının ucunda, dışbükey bir toynak taşıyan kısa bir parmak kemiği vardı. Thewissen ve meslektaşları Ambulocetus’un günümüz memeli deniz hayvanlarından farklı olarak, büyük olasılıkla paletleri olmayan uzun bir kuyruğa sahip olduğunu ileri sürmüşlerdir. İnsan, ucunda toynak bulunan bir ayağa sahip arka üyelerin ya da karada yürümek için tasarlanmış güçlü herhangi bir çeşit ön ve arka üyenin, bir balinada ne işe yaradığını merak ediyor.
Ambulocetus fosilinin, yaprak izleri ve içinde bol miktarda deniz karındanbacaklısı olan Turritella bulunan bir balçık ve çamur taşı yatağında bulunduğu bildirilmiştir. Bu da, bu canlının, deniz kıyısına yakın yerlerde yaşayan, belki kara hayvan veveya bitkileriyle beslenen ve belki de yumuşakçalar ve karındanbacaklılar ile beslenebilmek için sığ denizlerde yiyecek arayan bir canlı olabileceğini göstermektedir. Thewissen ve meslektaşları, fosil yataklarının Alt–Orta Eosen zamanına ait olduğunu ve tabakaların Pakicetus’un bulunduğu tabakalardan yaklaşık 120 metre daha yüksekte olduğunu bildirmişlerdir. Thewissen ve diğerlerinin makalesi hakkında yazdığı yorumda Berta, Pakistan’daki Ambulocetus’un bulunduğu tortular için 52 milyon yıllık bir yaş vermektedir. Thewissen ve meslektaşları da yazılarında, “en eski deniz memelisi” olarak sözünü ettikleri Pakicetus’un yaşı için 52 milyon yıllık bir yaştan bahsetmektedirler. Büyük ön üyeler ve toynaklı arka üyelere sahip olan Ambulocetus, Pakicetus’tan 120 m. yukarıdaki tabakalarda bulunmuştur. Bu nedenle Ambulocetus Pakicetus’tan daha yaşlı olamaz. Pakicetus, en yaşlı memeli deniz hayvanı olarak nitelendirilmiştir. Yine de Ambulocetus’un, balinaların evrimindeki geçiş hareket biçimlerini belgelediği söylenmektedir. Kafanız mı karıştı? Bizim de öyle. Ambulocetus’un dişlerinin, evrimcilerin eski balinalar ya da balinaların ataları olduğuna inandıkları diğer archeocete’lerin dişlerine benzediği bildirilmiştir. Ancak archeocete’lerin dişleri, kurt benzeri etçil memeliler olduklarına inanılan mesonychid toynaklıların dişlerine çok benzediği için, sadece dişlerinden tanınan iki archeocete Gandakasia ve Ichthylestes, ilk önce mesonychid’ler olarak sınıflandırılmışlardı.102
Balinalar konusunda uzman olan Rus G. A. Mchedlidze, Archeoceti’nin kara memelileri ve tipik Cetacea (deniz memelileri) arasında bir geçiş seviyesi durumunda bulunduğunu iddia ederken, Archeoceti ve günümüz Cetacea’sı arasındaki soy oluşsal ilişkideki problemin oldukça tartışmalı bir konu olduğunu söylemektedir. Mchedlidze, birtakım yazarların Archeoceti’yi, tipik Cetacea ile ortak hiçbir özelliği olmayan, tümüyle farklı bir grup olarak düşündüklerini bildirmiştir.103 Eğer bu düşünce doğruysa, eski balinalar oldukları varsayılan archeocete’ler aslında balina değillerdi ve balinalara (cetacea’lara) ata olmamışlardı.
Ambulocetus’a benzeyen canlıların fosilleri için memelilerle ilgili metinlerin araştırılması, Ambulocetus’a çok benzeyen tek bir canlı bulamamış ama morslara öncülük eden nesli tükenmiş bir deniz etçili olan Allodesmus’un Ambulocetus’a biraz benzediğini ortaya koymuştur.104
Eğer Robert Carroll’un Omurgalı Paleontoloji ve Evrim adlı çok kalın kitabında yer alan şu inanılmaz ifadeleri okursak, Thewissen ve meslektaşlarının Ambulocetus’u bir “balina” olarak isimlendirmeye cesaret etmelerine çok fazla şaşırmayacağız: “Görünüş bakımından çok büyük farklılıklar olmasına karşın, soy oluşsal sınıflandırma açısından mesonychid’leri Cetacea içinde sınıflandırmak mantıklıdır.”105 Sihirbazlığa bakın! Bu kurt benzeri canlılar, artık balina oldular! Kim demiş evrimcilerin geçiş formları yok diye? Bir kurdu, bir balina olarak isimlendirebilen bir kişi, “geçiş formları” bulmakta da hiçbir zorluk çekmeyecektir.
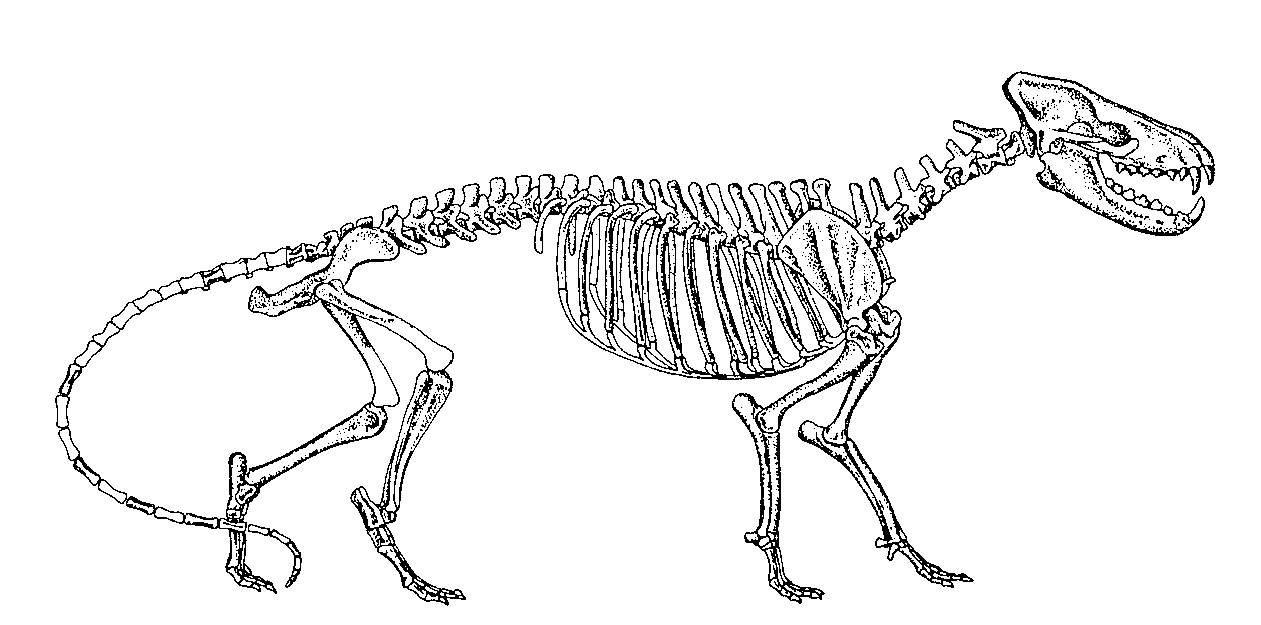
Şekil 17. Mesonyx. Kuzey Amerika’nın Eosen katmanlarından çıkarılmış etçil mesonychidiskeleti. Journal of the Academy of National Sciencesof Philadelphia 9:155 (1888).
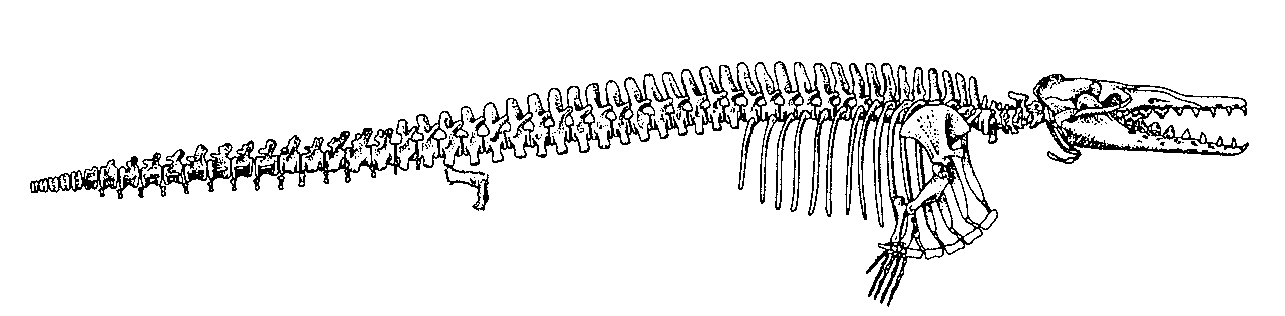
Şekil 18. Uzunluğu 24 metreden daha fazla olan Basilosaurus. Balinaların atası olarak önerilen canlılardan biri. Kellogg’dan, Carnegie Institute Washington Publication 482:1-366 (1936).
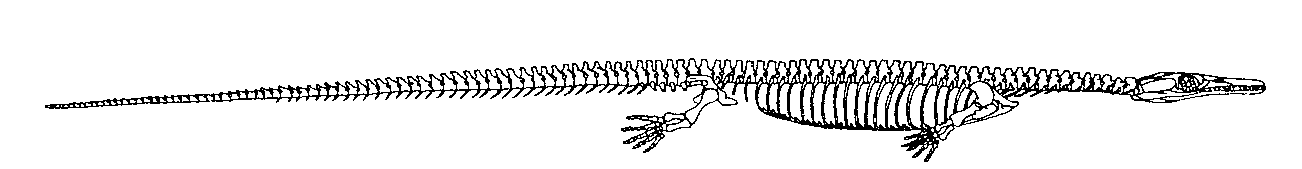
Şekil 19. İsviçre’nin Orta Trias katmanları arasında bulunan denizel sürüngen Askeptosaurus. Yaklaşık olarak 2 metre uzunluğunda. Kuhn Schnyder’den, Neujahrsblatt Naturf.
Ges. Zuerich 176:1-119 (1974).
Bu dizinin en son bölümü (bu kitap yayınlanmadan önceki), Gingerich, Raza, Arif, Anwar ve Zhore’nin balinalar ile kara memelileri arası bir ara seviye olduğunu iddia ettikleri sözde archaeocete olan başka bir fosil keşfettikleri konusundaki bir rapordur.106 Onlar bu canlıyı Rodhocetus kasrani olarak isimlendirdiler. Bu fosili, Pakistan’ın Punjab ilinin güneybatısında buldular ve fosilin 46-47 milyon yıl yaşında olduğuna inanmaktadırlar. Bu canlının, kendi ağırlığını karada taşıyabildiğini gösterdiğine inandıkları bazı özelliklerini tarif ettiler. Ayrıca bu araştırmacılar, bu canlının iyi bir yüzücü olduğunu gösteren niteliklere sahip olduğuna ve dolayısıyla kara memelileri ile deniz memelileri arasında bir ara seviye oluşturduğuna inanmaktadırlar.
Bu canlılar gerçekten de, evrimsel ataları kurt benzeri, torunları da balinalar ve yunus balıkları olan ara seviyeler miydi? Yoksa Mchedlidze’nin de inandığı (ve başkalarının da sözünü ettiği) gibi, sıradan balinalarla hiçbir ortak özelliği bulunmayan farklı bir grup muydu? Yaratılışçı bilim adamları, evrimci Mchedlidze ve onun evrimci meslektaşları şu noktada hemfikirdirler: bu canlılar, yüzgeç ayaklılar (normal ve büyük fok balıkları ile moslar) ile sirenia’larda (denizayıları ve dugonglarda) olduğu gibi karışık özellikler taşımakta, fosil kaydında hiçbir geçiş formu izi olmaksızın, tümüyle gelişmiş biçimde ortaya çıkmakta ve balinalar ya da yunus balıklarının sözde atalarıyla hiçbir ilişkileri bulunmamaktadır.
Günümüzde iki farklı balina türü vardır: dişli balinalar olan odontocete’ler (Odontoceti alt takımı) ve baleen balinaları olan mysticete’ler (Mysticeti alttakımı). Odontoceti’lerin arasında, katil balinalar, deniz gergedanları (bu canlının erkeğinde, üst çenesinden uzayan helezon biçiminde bir fildişi vardır), ispermeçet balinaları ve yunus balıkları vardır. Günümüz balinalarının en büyükleri olan baleen balinalarının arasında, gerçek balinagiller, Balaena Mysticetus’u, sırt-yüzgeçli ya da norqual balinalar ve uzunluk olarak 30 metreye ve ağırlık olarak da 150 tona kadar ulaşabilen mavi balinalar yer almaktadır.
Balinaların yaşam ortamları ve yaşam biçimleri, karada yaşayan hayvanlarınkinden çok farklıdır. Bu farklılıklarla ve birçok değişik yaşam biçimiyle başa çıkmak için balinalar, çok sayıda, inanılmaz derecede özelleşmiş üyelerle donatılmışlardır.107 Evrimciler, ihtiyaç duyulan şey ne olursa ve ne kadar karmaşık ve alışılmadık olursa olsun, rastlantısal genetik hataların, gerekli olan yapıları mükemmel bir şekilde ayarlanmış olarak yapabildiğine inanmak zorunda kalmaktadırlar.
Balinalar, hem ahtapotlarla hem de karideslerin bol miktarda bulunduğu yerlerde karideslerle beslenmek için, büyük derinliklere dalmak zorundadır. Sıradan hayvanlar bunu yapamazlar. Önceden tartışılan Basilosaurus, Pakicetus, Ambulocetus, Rodhocetus gibi sözde ara seviyelerin hiçbiri bunu yapmak için donatılmamıştır. Gagalı yunus balıkları yaklaşık 365 metre derinliğe rahatlıkla dalabilirler. Gagalı balinalar 490 metreden daha derinlere dalabilirler. Dişli balinaların en büyükleri olan ispermeçet balinaları (yaklaşık olarak 20 metre boyunda ve 55 ton ağırlığındadırlar) rahatça 900 metre ve hatta 3 km. derinliğe dalabilirler.
Bu kadar çok derinliklerdeki büyük basınca dayanabilmesi için, balina kulağının çok daha fazla damarlanması ve bunun yanında balinanın işitme organları ile kafatasının çok özel şekilde ayarlanmış olması gerekmektedir. Sadece 900 metre derinlikte bile basınç deniz seviyesindekinin 100 katına çıkmaktadır. İspermeçet balinasında, suyun derinliğine ve sıcaklığına göre yüzdürücülüğün ayarlanabilmesi için değişim gösteren yüzlerce litrelik ispermeçet yağı içeren büyük bir kapalı bölme vardır. Bu balina dalmadan önce, kanında, kaslarında ve akciğerlerinde oksijen depolamak amacıyla on dakikalık bir nefes alma alıştırması yapar. Bu balinaların kanı, insan kanındaki hemoglobinden %50 daha fazla hemoglobin içerir ve insanlar enerji harcarlarken aldıkları havanın sadece %10-20’sini kullanırlarken bu balinalar %80-90’ını kullanabilirler. Bir dalış süresinde, oksijenin %41’i kandan ve %50’si de kaslar ile dokulardan elde edilirken, akciğerlerden yalnızca %9’u elde edilir. Balinalarda, bu inanılmaz dalışlar için başka birçok değişiklik de gereklidir. Bu balinalar bu derin dalışlarında, dev ahtapotların yanı sıra binlerce mürekkepbalığı tüketmektedirler (bir ispermeçet balinasının midesinde 28,000 tane bulunmuştur).
Dişli balinalar, derinliklerde, karanlıkta “görebilmek” için bir ekolokasyon sistemi, diğer bir deyişle sonar sistemiyle donatılmışlardır. Bu balinaların, 95 km uzaklıktan su altına gönderilen sesleri duyabildikleri kaydedilmiştir. Ayrıca bu balinalar su altında şarkı söylerler ve oldukça geniş bir repertuara da sahiptirler.
Balina yavruları su altında doğarlar. Eğer bu yavrular, normalde insan bebeklerinin doğduğu biçimde baş aşağı doğmuş olsalardı hayatta kalamayacaklardı. Tüm balinaların önce kuyrukları doğar. Balina yavruları su altında emzirilmelidirler. Eğer bildiğimiz yolla emzirilmek zorunda kalmış olsalardı ya boğulacaklar ya da açlıktan öleceklerdi. Ama durum böyle değildir. Anne balinanın meme bezleri, sütün, bu basınç altında, su yüzeyinde iki metre yükseklikte bir fıskiye oluşturabilecek kadar şiddetli biçimde yavrunun ağzına fışkırtılmasını mümkün kılacak şekilde donatılmıştır. Anne balinanın sütü %42 yağ ve %12 protein içerir; bu oran insanda %4.4 yağ ve %1 protein şeklindedir. Yavru bir mavi balina günlük 90 kg. süt içer ve her gün 80 kg. ağırlaşır.
Bazı evrimciler süzgeçli ya da baleen balinalarının (Mysticeti) ve dişli balinaların (Odontoceti) farklı kökenlere sahip olduklarına inanırken bazıları da bunların ortak bir archaeocete atadan evrimleştiklerine inanmaktadır. Odontocete’ler ve mysticete’ler fosil kaydında ilk ortaya çıktıklarında, yaratılışın öngördüğü gibi zaten tamamıyla birbirlerinden farklıydılar.108 Baleen balinaları, katılaşmış deri çıkıntısından oluşan ve çapraz paralel sıralar halinde ağızdan aşağıya doğru uzanan bir balina çubuğuna ya da baleen’e sahiptiler. Bu balinalar, su yüzeyinde yüzen mikroskobik organizmalar olan planktonları çıkarıp onlarla beslenebilmek için büyük miktarda deniz suyunu süzerler. Dev mavi balinalar, ağız, boğaz ve göğüslerinin altındaki devasa soluk keselerinden faydalanarak bir kerede yetmiş tonluk suyu yutabilirler.
Açıkça görülmektedir ki deniz memelilerinin kökenleriyle ilgili kanıtların büyük çoğunluğu yaratılışın tarafındadır. Dört ayaklı kıllı bir memelinin suya yönelip kademeli ve çok uzun süreçler sonunda, binlerce rastlantısal genetik hata yoluyla balinaları, yunus balıklarını, deniz ayılarını, normal ve büyük fokları, morsları ve diğer deniz memelilerini ortaya çıkardığına inanmak, materyalist felsefenin daha önce yasakladığı mucizelere inanmayı gerektirmektedir. Bu bilinçsiz deneme yanılma yöntemi, sözde balinaların onlarsız yaşamayacağı ve oluşumlarının ilk evrelerinde tamamen yararsız ve aslında zararlı olacağı çok yüksek özelleşmiş karmaşık organları ve yapıları güya ortaya çıkarmıştır. Evrim kuramı, inanılmaz bir inançtır.
Özet
Daha önceden de belirtildiği gibi, sürüngenler ile memeliler arasında çok sayıda temel farklılıklar bulunmaktadır. Memelilerdeki bazı organlar, Korti organı gibi, sürüngenlerde hiçbir benzer yapısı bulunmayan ve yoktanvar edilmiş olmak zorunda olan inanılmaz derecede karmaşık yapılardır. Memelilerdeki üreme sistemi, sürüngenlerdekinden temelde çok farklıdır. Yalnızca memelilerin meme bezleri vardır. Sadece memelilerde diyafram vardır ve sürüngenlerde, bu yapının türeyebileceği nitelikte hiçbir benzer yapı bulunmamaktadır. Balina ve yarasalardaki karmaşık ekolokasyon sisteminin kökeni evrimsel bir açıklamaya yer vermemektedir.
Bu konuda belirtildiği ve iyice belgelendiği gibi, fosil kaydında başlıca türler arasında bulunan boşluklar sistematik ve genel olarak büyük boşluklardır. Geçiş formları, özellikle de yarasalar gibi ileri derecede özelleşmiş memelilere ata oldukları hayal edilenlere ait geçiş formları fosil kaydında bulunmamıştır. Bizler, Gould ve Eldredge’nin şu ifadelerine fazlasıyla katılıyoruz:
Temel morfolojik tasarımların arasındaki evrimsel geçişin yüksek seviyelerinde kademeli değişim, çoğu batılı evrimcinin “resmi” kanısı olmasına rağmen hep zor durumda kalmıştır. Baupläne arası düzgün ara seviyelerin bir araya getirilmesi düşünsel denemelerde bile neredeyse imkansızdır; bu düzgün ara seviyeler için fosil kaydında kesinlikle hiçbir kanıt bulunmamaktadır (Archaeopteryx gibi tuhaf karışımlar hesaba katılamaz).109
7. BÖLÜM
İnsan, Homo sapiens, memelilerin otuz iki takımından biri olan Primates takımında yer almaktadır. Yaşayan primatlar arasında, yarı maymunlar (lemurlar, lorisler ve tarsierler), Yeni Dünya maymunları, Eski Dünya maymunları ve kuyruksuz maymunlar (gibonlar, siyamanglar, orangutanlar, goriller ve şempanzeler) yer almaktadır. İnsan ve bu canlılar Primates takımında bir arada sınıflandırılmıştır; çünkü belirli nitelikleri ortak olarak paylaşmaktadırlar. Örneğin tüm primatlar kavrayıcı ellere, güçlü bir görme ve işitme yeteneğine, nispeten az bir koklama yeteneğine ve nispeten büyük beyinlere sahiptirler (insanların kafatası hacimlerinin ortalaması, herhangi yaşayan bir kuyruksuz maymununkinin üç katıdır). Evrimciler bunun doğal bir gruplama meydana getirdiğini düşünürler; çünkü onlara göre bu benzerlikler ortak bir atadan gelmelerinden dolayı vardır. Ancak, embriyolojik, morfolojik ve genetik kanıtların, farklı hayvanlarda benzer (homolog) yapıların varlığının ortak atadan kalıtım nedeniyle oluştuğu fikrine karşı olduğu bir sonraki bölümde görülecektir. Yaratılışçı bilim adamları bu benzerliklerin var oluş nedeninin, bu canlıların yaşam biçimlerinin bu nitelikleri gerektirmesi olduğunu varsaymaktadırlar. Maymunlar, kuyruksuz maymunlar ve yarı maymunların ağaçlarla ilgili etkinlikleri, kavrayıcı elleri olmasını gerektirir. İnsanların ve bu canlıların, yiyecekleri ve diğer nesneleri ustalıkla kullanabilmesi için kavrayıcı ellere ihtiyaçları vardır. İnsanlar ve diğer primatlar burunlarını yere koyarak av peşine düşmedikleri için, güçlü bir koku hissine ihtiyaçları yoktur fakat, pek çok etkinliklerinde güçlü görme ve işitme hislerine ihtiyaçları vardır. Yaşam biçimleri, daha yüksek bir zeka seviyesine dolayısıyla da daha büyük ve daha etkin bir beyne sahip olmalarını gerektirir. Taksonominin babası ve bir yaratılışçı olan Carolus Linnaeus, insan ve diğer canlıları Primates takımında toplayan ilk kişidir; elbette ki bunu evrimsel bir anlam ifade etmeden yapmıştır.
|
Takım: Primates |
||
|
Alt takım: Prosimii |
Alt takım: Antropoyitea |
|
|
Infra-takım: Lemuriformes |
Infra-takım: Platyrrhine |
Infra-takım: Catarrhini |
|
Üst familya: Lemuroidea |
Familya: Cebidae |
Familya: Cerropithecidae |
|
Familya: Lemuridae |
Yeni Dünya Maymunları |
Eski Dünya Maymunları |
|
Lemurlar |
Üst familya: Hominoidea |
|
|
Üst familya: Lorisoidea |
Familya: Pongidae |
|
|
Familya: Lorisidae |
Pan: Şempanze |
|
|
Lorisler |
Gorilla: Goril |
|
|
Infra-takım: Tarsiiformes |
Pongo: Orangutan |
|
|
Familya: Tarsiidae |
Familya: Hominidae |
|
|
Tarsierler |
Homo: İnsan |
|
Şekil 20. Günümüz Primatlarının Sınıflandırılması
İnsanın kuyruksuz maymunlar dahil diğer tüm canlılardan esasen farklı olduğu, yaratılışçı bilim adamları ve pek çok insan için inkar edilemez gibi görünmektedir. Evrimciler insanın, kuyruksuz maymunlardan sadece biraz daha yüksek bir derecede evrim geçirdiğini tartışsalar da bu kişiler kuyruksuz maymun ile insan arasında derin farkların var olduğunun farkındadırlar. Cartmill, Pilbeam ve Isaac bunu şu şekilde açıklamışlardır:
İnsanlığın doğaüstü kökeni konusundaki geleneksel Batı fikirlerinin zihinsel saygınlığını yitirdiği Darvinci devriminden itibaren, bilim adamları insan türlerinin ortaya çıkışını doğal nedenlerin bir sonucu olarak açıklamak için çok uğraşmaktadırlar. Ama onlar genellikle kendi görevlerini, insanların diğer hayvanlardan bu kadar çok ve bu kadar önemli bir şekilde nasıl farklılaştıklarını açıklamak olarak görmektedirler. İnsan evrimi üzerinde çalışan bilim adamları, çözmeleri gereken sorunlarını bu Darwin öncesi kalıcı tanımı olarak kabul ederek kendilerine, doğanın her yerinde işleyen nedenlerin, Homo sapiens’in doğadaki başka hiçbir şeye benzemediği gibi radikal bir sonuca nasıl vardığını açıklamak gibi mantığa aykırı görünen bir işi yüklemişlerdir.1
Evet; gerçekte insan, kuyruksuz maymunlar dahil radikal bir şekilde doğadaki başka hiçbir şeye benzememektedir ve Darwin’den bu yana evrimciler, insanın atasına dair hayvansal bir kanıt keşfetme girişimlerinde çok rahatsız edici biçimde hayal kırıklığına uğramaktadırlar. Holmquist, Mujamoto ve Goodman “İleri-Primat Soyoluşu: Niçin Karar Veremiyoruz?” adlı makalelerinde şöyle demektedirler:
Günümüzde, onlarca yıl morfolojik ve immünolojik protein ile nükleik asit sırasına ait verilerin toparlanıp bütün haline getirilmesine ve bu verilerin analizlerinin anlaşılması ve yorumlanması için çok sayıda mantıklı kuramsal model ortaya atılmasına rağmen, ileri primatların soyağacındaki dallanmanın doğru sıralandırılması ya da ileri primatlar arasındaki evrimin farklı hızlardaki oranı konusunda kesin bir fikir birliği yoktur. Burada söz konusu olan, üç farklı köksüz soyoluş ağacı...2
Uzun yıllar boyunca binlerce biyokimyacı, immünolog, paleoantropolog, biyolog ve diğer bilim adamları tarafından yıllarca yapılan araştırmalar sonucu ortaya çıkan tüm veriler göz önüne alındığında, bugün, insanın evrimiyle ilgili olarak niçin hiçbir fikir birliği bulunmamaktadır? Bunun nedeni, tam onların da itiraf ettikleri gibi evrimsel soyoluş ağacının hiçbir kökünün olmamasıdır. Ayrıca bu ağacın ne bir gövdesi, ne de dalları vardır. Ağaçta var olan tek şey, dal uçlarıdır.
Paleoantropolog Bernard Wood rahatsızlığını şu şekilde dile getirmektedir:
Kendi cinsimiz olan Homo’nun bilinen en erken temsilcilerinin sınıflandırılmalarının ve soyoluş ilişkilerinin karanlıkta kalması dikkate değerdir. Fosillerin yeniden yorumlanması ve kesin tarihlendirmelerinin yapılması konusundaki teknik gelişmeler, Homo habilis’in australopithecine’lerin yerini aldığı ve daha sonra H. erectus’tan H. sapiens’e evrimleştiği basit ve tek dallı bir insan evrimi modelini savunulamaz hale getirmiş fakat başka net bir görüş birliği henüz ortaya çıkmamıştır.3
Jerold Lowenstein ve Adrienne Zihlman, insanın, beyninin genişlemeden önce de iki ayak üzerinde yürüdüğü şeklinde fosil kaydını yorumlamaya dayanan inançlarını duyurduktan sonra şöyle demektedirler:
Fakat evrimsel nesillerin tanımlamalarında, fosil kaydına ve anatomiye güvenilemez. Şimdiye kadar paleontologlar, yine de bunu yapmakta ısrar etmektedirler. Onlar, ortak olarak paylaşılan ya da paylaşılmayan “ilkel” ve “türemiş” özellikler (çoğunlukla dişler ve kemikler) temelinde inşa edilmiş canlı ve fosil primatlar ağacının yer aldığı, kladistik sınıflandırma olarak isimlendirilen bir yöntem altında bir araya toplanmışlardır. Paylaşılan ortak ilkel nitelikler, ortak bir atadan geldikleri için paylaşılmaktadır; paylaşılmayan türemiş nitelikler ise farklı evrimsel yolları açığa çıkarmaktadırlar. Pek çok paleontologun neredeyse dini hevesle savundukları evrimsel ağaçların inşasındaki bu yaklaşımın öznelliği şu sonuçlarla gösterilmiştir: onların, fikir birliğine vardıkları tek bir familya ağacı bile yoktur. Aksine, kladistik sınıflandırma yapan paleontologlar, canlı ve soyu tükenmiş hominoidlere dair neredeyse akla gelebilecek tüm permütasyon ve kombinasyonlarını ileri sürmüşlerdir.4
Görünüşe göre Lowenstein ve Zihlman makalelerinin bir önceki kısmında aşağıdaki itirafları yapmış olmalarına rağmen, yargılarındaki öznelliği fark etmemişlerdir:
Hem kuyruksuz maymunların hem de insanın atası olan bizim en eski atamızın resmini canlandırmak, hayallerimizi birbirine karıştırır. Bu ata, ne kuyruksuz maymun ile insan anatomisinde, ne de fosil kaydında belli değildir; fakat yalnızca hücre genomlarının görünmez dünyasında açık bir şekilde bellidirler.5
Lowenstein ve Zihlman diğer paleoantropologların da itiraf ettikleri gibi şunu açığa vurmaktadırlar:
Geçtiğimiz yüzyılda her yeni hominid ya da hominoid kaşifi, buldukları şeyi olası bir insan atası adayı olarak göstermiştir.6
Üstelik, keşfedilen hemen hemen her kemik kırıntısına yeni bir tür adı verilmektedir. Bir paleoantropologun, insanın atalığına giden gerçek yolu gösteren bir fosil bulunmadıkça, özellikle de bu fosil için bu alandaki en eski fosil olduğu iddia edilmedikçe, ne başkasının adımlarıyla zar zor yürümesi zevkli olur ne de paleoantropolojide şöhret kazanabilir. Ve insanın atası araştıran bilimin başka hiçbir alanı, bu kadar ideolojiyle yoğun değildir. Bu durum, insanların nihai sonuçlarının ve her türlü algılarının kendi dünya görüşleriyle belirlendiği bu bilim dalında kaçınılmazdır.
Evrimciler ilk primatların, bir tartışma konusu olmasına rağmen, ya lemur benzeri bir canlıya (adapid’ler) ya da “yarı maymunların modern görünüşü” olarak sözü edilen tarsier benzeri bir canlıya (omomyid’ler) benzediğini varsaymaktadırlar. Bu canlıların fosilleri Kuzey Amerika, Avrupa ve Asya’nın Eosen kayaları arasında bulunmuştur. Çok yakın zamana kadar, Paleosen kayaları arasında fosilleri bulunan üç canlı, Phenacolemur, Ignacius ve Plesiadapis’in, yarı maymunların erken ataları olabileceği düşünülüyordu. Araştırmalar artık bu canlıların primatlarla akraba olmadıklarını fakat, bir günümüz calugo’su ya da “uçan lemuru” (bu canlı ne bir lemurdur ne de uçabilir; sadece havada süzülür) olan Cynocephalus’a benzediğini göstermiştir.7 Bir paleoantropologun üzülerek söylediği gibi: “Bildiğimiz erken primat evrimi, pencereden süzülüp gitmiştir diyebiliriz.” Bu gerçekler, yarı maymunlar ve önerilen ataları arasında büyük bir boşluk oluşturmuştur.
Primatların böcekçil bir atadan evrimleştiği ileri sürülse de, primatları böcekçillere bağlayan bir geçiş formu dizisi bulunmamaktadır. Primatlar dalında dünyanın lider uzmanlarından biri olan Elwyn Simons şöyle kabul etmek zorunda kalmıştır: “Günümüz bulgularına rağmen Primatlar takımının kökeninin bulunduğu yer ve zaman, sırlarla örtülü olarak kalmıştır.”8 Romer’in söylediğine göre ilk lemurlar “görünüşe göre, bilinmeyen bir bölgenin göçmenleri olarak” ortaya çıkmışlardır.9 Romer bunu, paleontologların, lemurların nasıl ortaya çıktıklarını fosil kaydıyla söyleyememeleri nedeniyle söylemek zorunda kalmıştır. Kelso şöyle demiştir:
…böcekçilden primata dönüşüm, fosil kaydında belgelenmemiştir. Dönüşüm hakkındaki bilginin temeli, canlı formlardan yapılan çıkarımlardan kaynaklanmaktadır.10
Bu durum bu yayınlardan sonra da değişmemiştir; çünkü Carroll şöyle demektedir:
Primatların, daha ilkel gerçek plasentalılar arasındaki kesin kökeni belirlenmemiştir… İlk primatlar ve diğer herhangi bir takımın ilk üyeleri arasında, ortak belirgin hiçbir türemiş nitelik gösterilmemiştir.11
Öyleyse görebiliriz ki daha başlangıçta, tüm primat takımının kökeni fosil kaydınca belirlenememiştir. Eğer primatlar evrimleşmişlerse, bizi böcekçil atalara doğru yönlendiren bir dizi geçiş formu bulunmalıdır ama böyle geçiş formları bulunmamaktadır. Tabi ki yaratılışçıların, kayıtlardan göstermesini beklediği şey de budur.
Evrimcilerin primatların atası olarak önerdikleri belirgin böcekçil, sivri sincapçıktır. Bu karar, 1920’de, Wilfred Le Gros Clark’ın, Asya sivri sincapçığı olan Tupaia’yı araştırması esnasında alınmıştır. Le Gros Clark, primatlar ve sivri sincapçıklar arasında çok sayıda benzerlik görebildiğini sanmıştır. Ancak, geçtiğimiz birkaç on yıl süresince yapılan çalışmalar, Le Gros Clark’ın gerçekten yanıldığını ortaya çıkarmıştır. C. B. G. Campbell iddia edilen bu ilişki hakkında daha 1966’da yazdığı eleştiride şöyle demektedir:
Tupai’giller ve primatlar arasında hiçbir yakın ilişkinin bulunma olasılığının olmadığını gösteren çok sayıda günümüz çalışmasını göstermeye çalıştım.12
Campbell, sivri sincapçık– lemur– tarsier– kuyruksuz maymun– insan sıralamasının kendine özgü çekiciliğinin, bu sıralamanın kabul edilmesinden büyük oranda sorumlu olduğunu düşünmüştür. Bu sıralamanın kabul edilmesindeki diğer bir etken de, kuşkusuz Le Gros Clark’ın antropolojideki yetkili durumuydu. Uzman kişi konuştuğunda, diğerleri ağzını açamaz.
Daha yakın zamanda R. D. Martin sivri sincapçıklar ve primatların annelik davranışları konusunda araştırmalar yapmış ve “sivri sincapçığın, insan ataları listesinde bulunmadığı” sonucuna varmıştır.13 Martin, hepsinin zahmetli annelik yaptığı dişi primatların tersine, dişi sivri sincapçıkların, yavrularını emzirdiği süre içerisinde yuvayı her kırk sekiz saatte bir sadece on dakikalığına ziyaret ettiğini bulmuştur. Bu, onun yavrularına gösterdiği tek bakımdır. Üstelik, sivri sincapçığın yavrusuna verdiği sütteki yağ oranı %25 iken, bu oran primatlarda genellikle %1 ile %3 arasında olmakta ve asla %5’i aşmamaktadır. Martin ayrıca W. P. Luckett tarafından yayına hazırlanan ve yakın zaman önce yayınlanan bir kitapta sivri sincapçığa verilen geniş kapsamlı değerlendirmenin, sivri sincapçıklar ile primatlar arasında belirli herhangi bir bağ olabileceğine pek olanak vermemektedir.
Yarı maymunlar ile sivri sincapçıklar arasında hiçbir bağlantı bulunmadığına dair morfolojik ve fizyolojik kanıtlar, çoğalmaya devam etmektedir. Martin, primatların kökeni konusunda yazdığı 1990 yayınında şunu vurgulamaktadır: “Bugün, sivri sincapçıkların primatlarla akraba olmadıklarına dair çok sayıda kanıt vardır.”14

Şekil 21. Filipin Adalarına özgü bir tarsier. Zoological Society of San Diego’dan 1995. İzinle kullanılmıştır.
Çin’in Jiangsu ilinin güneyindeki Shanghuang’a yakın bir yerde olduğuna inanılan çatlak dolgusunun içinde, heyecan verici bir fosil mezarlığının keşfi sonucunda, pek çok memeli fosilinin yanı sıra, her biri farklı bir soyu temsil eden beş yeni “ilkel” primat türü ortaya çıkartılmıştır.15 Primatlar, adapid’leri, omomyid’leri ve modern bir tarsierden hemen hemen ayırt edilemeyen bir canlı fosilini içerirler. Bu yüzden canlı bir tarsierin bugün, “canlı bir fosil” oluşturduğu söylenmektedir. Bu fosilleri keşfedenler, bu fosillerin yaklaşık kırk beş milyon yıl yaşında olduklarına inanmışlar ve böylece tarsierin yaşını önceki hesaplamalardan otuz milyon yıl daha öncesine atmışlardır. Üstelik bugün, adapid’ler ile omomyid’lerin günümüz tarsierleriyle aynı zamana ait oldukları bilindiği için, bu canlıların tarsierlerin ataları olmaları da mümkün değildir.
Kelso, Martin ve diğerlerinin de açıkladığı gibi, primatları sivri sincapçıklara (böcekçillere) ya da başka herhangi bir şeye bağlayan hiçbir geçiş formu fosili bulunmamaktadır. Campbell, Martin ve diğerleri, yapılan çalışmalarda, yaşayan sivri sincapçıkları primatlara bağlayan hiçbir kanıtın bulunmadığı gerçeğini belgelemişlerdir. Dolayısıyla ne günümüz dünyasında ne de geçmişin dünyasında, primatları başka herhangi bir canlıya bağlayan hiçbir kanıt yoktur. Öyleyse insanın evrimsel kökeni, gerçek deneysel bilimin ortaya çıkardığı kanıtlarla en başında geçersiz kılınmıştır. Bir grup olarak primatlar, diğer tüm canlılardan tümüyle bağımsız durumda bulunmaktadırlar.
Belirttiğimiz gibi antropoyitler, Yeni Dünya Maymunları (Platyrrhini) ve Eski Dünya maymunlarını (Catarrhini) içermektedirler. Evrim hikayesine göre antropoyitlerin, yarı maymun dalları olan adapid’ler veya omomyid’lerin birinden evrimleştiği varsayılmaktadır. Antropoyitlerin hangisinden evrimleştiği, hikayeyi anlatana bağlıdır. Ancak, maymunlar ya da antropoyitler ile yarı maymunlar arasında çok büyük bir boşluk vardır. Genellikle, geniş burunlara sahip olan Yeni Dünya Maymunları’nın, dar burunlu maymunlar olan Eski Dünya Maymunları’ndan farklı bir evrimsel kökeni olduğu varsayılmaktadır. Ancak, maymunları başka herhangi bir şeye bağlayan hiçbir geçiş formu fosili bulunmamaktadır. Antropolog Susan Cachel şöyle demektedir:
Elimizde var olan fosil kanıtları, yarı maymun–antropoyit dönüşümünü belgelememektedir... ve bu nedenle, günümüze kadar gelen antropoyitlerin morfolojileri üstün bir önem kazanmaktadır.16
Elizabeth Culotta şöyle demektedir:
Bir noktada ilkel primatlardan (yarı maymunlardan) bir grup ayrıldı ve antropoyitlerin oluşmasına neden oldu. Fakat bu atanın neye benzediğini hiç kimse kesin olarak söyleyememektedir. Çünkü, fosil kaydında ilkel ve gelişmiş formlar arasında büyük bir boşluk bulunmaktadır. Stony Brook’taki New York Eyalet Üniversitesi’nden John G. Fleagle şöyle demektedir: “Tüm primatları bir yere yığdıktan sonra, daima antropoyitleri diğerlerinden ayırt edebilirsiniz. Birbirlerinden öylesine farklıdırlar ki, nereden geldiklerini çözmek zordur.”... Fosil geçiş formlarının yokluğu, paleoantropologların, antropoyitlerin ataları olarak kendi beğendikleri adayları seçmelerini durduramamıştır...17
Yeni Dünya maymunları ile onların ataları oldukları varsayılan yarı maymunlar arasında hiçbir geçiş formu yoktur. Bu yüzden, Romer şöyle demektedir: “Ne yazık ki Güney Amerika maymunlarının fosil tarihleri konusunda çok az şey bilinmektedir.”18 Kelso şöyle demektedir:
Yeni Dünya maymunları olan Platyrrhinae’nin evrimsel geçmişi konusundaki ayrıntılar, kuşkusuz aydınlatıcı ve ilginç olacaktır; fakat ne yazık ki onlar hakkında çok az şey biliyoruz.19
Yeni Dünya maymunlarıyla ilgili olarak Carroll şöyle demektedir:
Platyrrhinae, kendi tarihi boyunca Güney ve Orta Amerika ile sınırlandırılmıştır. Bilinen en yaşlı fosil, Bolivya’nın Orta Oligosen’den Geç Oligosen’e kadar olan katmalarında bulunan, günümüz Cebidae familyasının bir üyesi olan Branisella’dır.20
R. D. Martin şöyle yazmaktadır:
Yeni Dünya maymunlarının, Güney ve Orta Amerika’daki bilinen fosil kaydı, açıkçası, pek çok açıdan oldukça hayal kırıcıdır. Kuzey Amerika’dan çıkarılan tüm sağlam primat fosilleri, yukarıda da gösterildiği gibi, Erken Üçüncü Dönem’in ya “lemurgiller”inin ya da “tarsiergiller”inin arasına dahil edilmektedir. Yeni Dünya maymunlarının kökeni, bu yüzden sır kalmaktadır; çünkü önemli fosillerin hepsi sadece Geç Oligosen ve Miyosen’den bilinmektedir, o aşamaya kadar bu fosiller onları modern formlara bağlayan birçok nitelik sergilemektedir. Dahası, istisnasız olarak, tarif edilen önemli fosiller, gerçek Yeni Dünya maymunlarına (Cebidae familyası) benzemektedir...21
Yeni Dünya maymunlarının, ilk ortaya çıktıkları anda birer maymun oldukları oldukça açıktır.
Catarrhine’ler, Eski Dünya maymunlarını (dar burunlu maymunları), kuyruksuz maymunları ve insanları kapsamaktadır. Bunların kökenleriyle ilgili olarak Simons şöyle demektedir: “Kelime olarak kullanılmasına rağmen gerçekte, fosil kaydında ‘protocatarrhine’ (ilkel catarrhine) diye bir şey bulunmamaktadır.”22 Simons, daha sonraki bir yayında şöyle demektedir:
Her iki kıtada da Eosen Çağı primat fosillerinin hiçbiri, insanın da dahil olduğu gelişmiş tüm canlı Eski Dünya maymunlarını kapsayan büyük catarrhine infra–takımının kabul edilebilir atası olarak görünmemektedir.23 Bu “ilkel” yarı maymunlar, o güne kadar yalnızca Kuzey Amerika ve Avrupa’da bulunabilmişlerdi.
A. J. Kelso şöyle yazmaktadır:
Açıkçası, Eski Dünya maymunlarının ortaya çıkışlarıyla ilgili fosil belgeleri, primatların genel evrimsel tasvirini kavramamızda bir anahtar olabilirdi fakat gerçekte, bu kayıtlar yoktur.24
R. D. Martin şöyle demektedir:
Yeni Dünya maymunlarının erken fosil kanıtları düş kırıklığına uğratıcı kanıtlar şeklinde açıklanabilse de, bu kanıtlar, Eski Dünya maymunlarınınkiyle karşılaştırıldıklarında oldukça zengindirler… Ayrıca fosil kaydı bize, Eski Dünya maymunlarının erken kökenleri hakkında çok az şey söyleyebilmektedir.25
Fosil kaydı, en erken primatlar olarak kabul edilen yarı maymunlar ile Yeni Dünya maymunları ya da Eski Dünya maymunları arası geçiş formları için kanıt sağlamakta başarısızlığa uğramıştır. Bu noktada, fosil kaydındaki başka büyük bir boşlukla karşı karşıya kalıyoruz. Geçiş formları, evrim kuramının onlara en çok ihtiyaç duyduğu yerlerde kayıptırlar.
Evrim merdivenindeki bir sonraki basamak, kuyruksuz maymun benzeri canlıların, maymun benzeri atalardan geldiği varsayılan kökenini içermektedir. Burada yine büyük bir boşlukla karşılaşmaktayız. Maymunlar ve kuyruksuz maymunlar arasında saptanabilir tek bir geçiş formu yoktur.
İnsanlar ve kuyruksuz maymunlar Hominoidae süper familyası içinde sınıflandırılmışlar ve bu tür canlıların tümü hominoid olarak nitelendirilmiştir. Hominoid’lerin evrimi konusunda önerilen bir tasarı, Şekil 22’de gösterilmiştir. Şekilden de anlaşılabileceği gibi, evrimciler arasındaki ortak karar, şempanze ve gorillerin, kuyruksuz maymunlar arasında bizim en yakın akrabalarımız olduğudur.
Şekil 22. Hominoid’ler için bir soy oluş tasarım önermesi.
Evrimciler, geçmişte bir zamanda, bir kuyruksuz maymun benzeri canlı popülasyonunun var olduğuna ve bu popülasyonun, kendi içinde alt popülasyonlara bölünüp, bir taraftan gorillerin atası olan bir popülasyonun, diğer taraftan da şempanze ve insanı oluşturabilecek biçimde kendi içinde yeniden bölündüğüne inanırlar. Kuyruksuz maymun ve insanın en son ortak atasının yaşadığı zaman, çok tartışılmaktadır. Güncel tahminler, hikayeyi anlatana bağlı olarak dört milyon yıl ile on dört milyon yıl arasında çeşitlilik göstermektedir. Belki de bu, bu konu hakkında aslında ne kadar az şey bilindiğinin bir göstergesidir. Paleontologlar, kuyruksuz maymun ve insanın varsayılan bu ortak atasını henüz bulmadıkları için, bu ata, tam olarak kuramsal bir canlı olarak kalmıştır.
Yukarıda da sözü edildiği gibi çoğu evrimci, şempanze (Pan) ve gorilin (Gorilla) insana, orangutandan (Pongo) daha yakın akraba olduğunu düşünmektedir. Bu yargının temeli, moleküler, biyokimyasal ve kromozomsal verilere dayanmaktadır. Örneğin çekirdeksel insan DNA’sı ile Afrika kuyruksuz maymunu (goril ve şempanze) melezleştirildiği zaman, %1,1 baz uyuşmazlığı oluşurken, yine insan DNA’sı ile orangutan DNA’sı melezleştirildiği zaman %2,4 baz uyuşmazlığı oluşmaktadır.26 Ancak, bu çeşit verilere dayandırılarak hazırlanan soy oluşun doğruluğu, Pittsburgh Üniversitesi’nde Antropoloji Profesörü olan Jeffrey Schwartz tarafından sorgulanmaktadır.
Schwartz, insanın, yaşayan hominoid’lerin yirmi altı niteliğinden, orangutanlarla yirmi altısını, şempanze ve gorillerle yalnız dokuzunu ve gibonlarla (Hylobates) ise beşini ortak olarak paylaştığını belirtmiştir.27 Schwartz’a göre insanın orangutanlarla paylaştığı fakat Afrika kuyruksuz maymunlarıyla paylaşmadığı özelliklerden bazıları şunlardır: en uzun saça, birbirinden uzak meme bezlerine, en uzun gebelik sürecine (insan ve orangutanda eşittir), kalın azıdişi minesine ve kısa taçlı yan dişlere sahip olma, adet dönemiyle sınırlandırılmamış bir çiftleşme ve adet dönemince oestriol seviyelerinin en yüksek olması. Schwartz bu verilerin, insanın orangutana, Afrika maymunlarından daha yakın akraba olduğunu gösterdiğini iddia etmektedir. Schwartz insanın, orangutandan çok, Afrika kuyruksuz maymunlarıyla sözde yakın bir bağa sahip olduğunu gösteren biyokimyasal ve moleküler verilerin büyük kısmının önemini reddetmekte ve geriye kalanları da, insanın, Afrika maymunundan çok orangutanla daha yakın akraba olduğunu gösterdiği şeklinde yorumlamaktadır.
Yaratılışçı bilim adamları, benzerliğin bir genetik ilişkinin varlığını gerektirmediğini iddia etmektedirler (insan ve hominoidlerin homologluğu, yani morfolojik özelliklerindeki benzerlikler bir sonraki bölümde tartışılacaktır). Bu yüzden de yaratılışçılar, evrimcilerin akrabalığı saptamak için kullandıkları benzerliklere göre oldukça çelişkili sonuçlara varabilmelerini ilginç bulmaktadırlar. Özellikle Schwartz’ın davası da böyle ilginçtir; çünkü Schwartz, insan ve kuyruksuz maymunlar arasındaki soy oluşsal akrabalığın saptanmasında biyokimyasal ve moleküler verilerin kullanımına karşı çıkmaktadır.
Geçiş formlarındaki sistematik yokluklar nedeniyle, fosil kaydına dayalı bir evrimsel soy oluş ağacı meydana getirme girişimlerinde büyük hayal kırıklığına uğrayan evrimciler son zamanlarda, akrabalığın saptanmasında moleküler verilerin kullanımı konusunda bas bas bağırmaktadırlar. Yaratılışçı bilim adamları, böyle verilere dayandırılan yargılara karşı çıkmakta ve Schwartz’ın sağladığı şekilde evrim çevresinden gelen yardımları memnuniyetle karşılamaktadırlar.
Sözde evrimsel akrabalığın saptanmasında, bu tür ilişkilerin anlamı, Benveniste ve Todaro’nun 1976’da yayınlanan çalışmalarıyla bazı yönlerden karıştırılmıştır.28 Daha önceden de sözü edildiği gibi, Benveniste ve Todaro tarafından yapılan DNA melezlemesi çalışmaları, Afrika kuyruksuz maymunlarının, yani goriller ve şempanzelerin, insana, Asya kuyruksuz maymunlarından, yani orangutanlar ve gibonlardan daha yakın akraba olduklarını göstermiştir. Ancak goril, şempanze, orangutan, gibon ve insan DNA’ları ile köpek maymununa (babon) ait C virüsünün DNA transkriptini melezleme araştırmaları, insan DNA’sının da, gibon ve orangutan DNA’larıyla aynı miktarda köpek maymunu virüsünün transkripti ile melezleştiğini ortaya çıkarmıştır. İnsan, gibon ve orangutan DNA’larının köpek maymununa ait C virüsünün DNA transkripti ile melezleşme oranları, goril ve şempanze DNA’larının melezleşme oranlarından daha azdı. Bu sonuçları sorgusuz kabul edersek, orangutan ve insan DNA’ları, benzer bir şekilde köpek maymununa ait C virüsü ile tepki gösterdiği fakat insan DNA’sının bu virüse gösterdiği tepkinin, goril ve şempanze DNA’larının bu virüse gösterdiği tepkiyle eşit olmadığı için, insanın, goril ve şempanzeden çok, orangutana daha yakın akraba olduğu gibi bir sonuç ortaya çıkmaktadır.
Böylesine bir yargı, Schwartz’ın ilgilendiği verilerle uyum içinde olmasına rağmen, yine de, diğer DNA melezlemesi çalışmalarından elde edilen sonuçlarla çelişkilidir ve pek çok evrimcinin ortak görüşüne göre insan, şempanze ve gorile, orangutana nazaran daha yakın akrabadır. Bu nedenle Benveniste ve Todaro şu fikri ileri sürmektedirler: köpek maymununa ait C virüsünün DNA transkriptlerinden elde edilen sonuçlar, insanın orangutan ve gibonla, goril ve şempanzeyle olduğundan daha yakın akraba olduğunu göstermemektedir fakat, bu benzerlikler ve farklılıkların var olmalarının gerçek nedeni, şempanze ve goril atalarının Afrika kökenli, insan atalarının ise Asya kökenli olmasıdır. Benveniste ve Todaro, insan atasının, goril ve şempanzenin atasından ayrıldıktan sonra Asya’ya göç ettiğini ve birkaç milyon yıl boyunca orada kaldığını ileri sürmüşlerdir. Bununla beraber insan evriminin büyük kısmının, köpek maymunları, goril ve şempanzelerin evi olan Afrika’nın tenha bir bölgesinde gerçekleştiğini ve insanın ya da insanın atalarının nispeten yakın zamanda Afrika’ya göç ettiğini varsaymışlardır. İnsanın, orangutanın ve gibonun virogenlerinin, Asya’da bulunmayan fakat Afrika’da yaygın olarak bulunan köpek maymununa ait C virüsüyle karşı karşıya gelmemesi, bu virogenlerin goril ve şempanzelerin virogenlerinden sözde daha fazla farklılaşmalarına neden oldu. Bu hayali hikaye anlatımı bizlere, evrim kuramının, bulunan veriler ne olursa olsun her şeyi içine alabilecek kadar esnekleştiğini bir kez daha hatırlatmaktadır. Bugün evrimcilerin çoğu insanın, Afrikalı bir kökene sahip olduğunu kabul etmektedir. Bizler de böyle verilerdeki benzerlikler ve farklılıkların bize, genetik akrabalıklar konusunda hiçbir şey anlatmadığı sonucuna varmaktayız.
Romer, şempanze ve gorilleri “antropoyit grubunun en ileri canlı üyeleri” olarak nitelendirmiştir. Peki Romer onların kökenleri hakkında ne söyleyebilir? Şöyle demektedir: “Bu ileri kuyruksuz maymunların ve varsayılan insanın atalarının fosil tarihi hakkındaki bilgimiz, bu seviyede, hayal kırıklığına uğratacak derecede zayıftır.”29 Bazı kişiler şempanze, goril ve orangutanların atalarının, Afrika, Avrupa ve Asya’da bulunan kuyruksuz maymun fosilleri olan Dryopithecus türleri arasında bulunabileceğini düşünmüştür.30 Neden orada bulunabileceği hiç de açık değildir.
İnsanın hayali kuyruksuz maymun benzeri atasıyla olan esas kökeni konusunda antropologlar ne söyleyebilirler? Pilbeam şöyle demektedir:
Pleistosen öncesi hominid atalarının, oldukça belirsiz bir biçimde olsa da, Dryopithecinae’nin içinde bir yerde kök saldığı genellikle varsayılır bir hal almıştır.31
Bir bilim adamının, bir şeyi “oldukça belirsiz bir biçimde varsaymak” zorunda kaldığı zaman çareyi, geçerli bilimsel yöntemlerde değil, bilimsel olmayan yöntemlerle kanıtlamakta aradığı açıktır. Paleoantropologlar, bir hayvanda, o hayvanın bir yandan şempanzelerin, gorillerin ve orangutanların atası olduğuna, diğer yandan da insan neslinin atası olduğuna karar verebilmelerini sağlayacak ne gibi tuhaf nitelikler bulabilirler ki?
Görünüşe göre Pilbeam, dryopithecine’lerin insanların atası oldukları genel varsayımına katılmamaktadır. Pilbeam, dryopithecine’lerin hominid’lerin atası olabilmeleri için fazla özelleştiğine, fazlasıyla maymunlaştığına dair inancını dile getirmiştir.32
Keşfedilmiş olan ilk iri kuyruksuz maymun olan Dryopithecus hakkındaki tartışmalar her zaman olduğu gibi bugün de canlıdır. Begum, Dryopithecus’un, büyük Afrika kuyruksuz maymunlarının çok yakın akrabası olduğunu iddia etmiştir33 (goriller ve şempanzeler büyük Afrika kuyruksuz maymunlarıdır, Asya’daki büyük kuyruksuz maymunlar olan orangutanlar Borneo ve Sumatra’da bulunmaktadırlar, küçük kuyruksuz maymunlar olan gibonlar ve siyamanglar, yani Hylobates güney Asya ve Doğu Hint Adaları’nda bulunurlar). Öte yandan Solà ve Köhler, İspanya’nın Orta Miyosen kayaları arasında bulunan ve sözde 10-12 milyon yıl yaşında olan yeni bir kafatası fosiline dayanarak, Dryopithecus’un, özellikle orangutanla bağlantılı olduğunu iddia etmektedirler.34 Orangutan, goril ya da şempanze ile benzerliği ne olursa olsun Dryopithecus, ne büyük kuyruksuz maymunların ne de insanların olası atası olabilmek için gerekli kabul edilen türemiş özelliklere (synapomorphies) sahipti.35
Peki, fosil kaydında günümüz kuyruksuz maymunlarına (goriller, şempanzeler, orangutanlar, gibonlar) önderlik eden geçiş formu kanıtları ve insanın varsayılan kuyruksuz maymun benzeri ataları olan australopithecine’lerin kanıtları nerede? Bu kanıtlar hiçbir yerde bulunmamaktadır. Watson şöyle demektedir:
Örneğin, günümüz kuyruksuz maymunları sanki bilinmeyen bir yerden aniden ortaya çıkmışlardır. Onlara ait ne bir dün ne de bir fosil kaydı bulunmaktadır. Ve günümüz insanlarının —iki ayaklı, çıplak, alet yapan, büyük beyinli canlılar— gerçek kökenleri de, eğer kendimize karşı dürüst olursak, aynı derecede sır dolu bir konudur.36
Ne yazık ki evrimcilerin çoğu Watson kadar açık sözlü değildir fakat, Watson’un ifadelerini doğrulamaktadır. Carroll şöyle demektedir:
Ne yazık ki Afrika’da, 4 ile 14 milyon yıl yaşları arasında, bilinen hiçbir hominoid fosili bulunmamaktadır. Afrika’ya ait kuyruksuz maymunlar olan Gorilla ve Pan’ın hiçbir fosil kaydı yoktur. Anatomik kanıtlara bakılırsa onlar, bundan yaklaşık 5 ilâ 14 milyon yıl önce insan atalarından ayrılmış olabilirler.37
Ayrıca R. D. Martin de bu gerçekleri doğrulamaktadır:
Öncelikle şu belirtilmelidir ki, aşağıda sıralanan tüm türler için sağlam fosil kalıntılarının varlığı bilinmektedir (genellikle primat fosil kaydı için oldukça alışılmadık bir durum), fakat 3,8 milyon yıl öncesinden kalan ve neye ait olduğu şüpheli birkaç parçacıktan başka hemen hemen hiçbir fosil kaydı yoktur. Bu nedenle, insan evriminin önceki devresi, kapalı bir sır perdesi arkasında kalmıştır ve hominid ile büyük kuyruksuz maymunların birbirlerinden ayrıldıkları zaman konusundaki görüş ne olursa olsun, fosil kaydında büyük, talihsiz bir boşluk vardır.38
“Bütün Primatlar Nereye Gitti?” başlıklı bir makalede Pat Shipman şunu anlatmaktadır:
Bugün, büyük kuyruksuz maymunların sadece üç cinsi ve dört türü vardır ve bunlar da hızla tükenen tropikal yağmur ormanları ya da ormanlık habitatlarla sınırlandırılmışlardır. Fakat, Miyosen çağından bilinen yaklaşık bir düzine cinste ve daha da fazla sayıda türde geniş vücutlu hominoid vardır. Bundan dolayı, antropologlar, canlı örneği olmayan pek çok eski kuyruksuz maymunla karşı karşıya gelmektedirler.39
Bir süre sonra yazar, primatların hareket yetenekleri konusunda uzman olan Mike Rose’den alıntı yapar:
Miyosen kuyruksuz maymunlarının postcranial kemiklerine baktığım zaman, birçok türün açık ve tutarlı bir modelini elde ediyorum. Fakat bu model, günümüz kuyruksuz maymunlarında gördüklerimize benzemiyor. Belki de hayatta kalmış olanları tuhaf olarak görmemiz gerekir.
Evrimcilerin günümüz kuyruksuz maymunlarını tuhaf olarak görmek istemelerinin tek nedeni, ilkel saydıkları kuyruksuz maymunlar içerisinde hiçbir uygun ata bulamamış olmalarıdır. Watson bize, günümüz kuyruksuz maymunlarının ve günümüz insanlarının kökenlerinin gizemli birer konu olduğunu söylemektedir. Martin bize, bundan dört milyon yıl önceki uzun süreli insan evriminin tam bir sır olduğunu söylemektedir. Belki de bu uzun bir dizi oluşturan “kayıp halkalar”, asla var olmadıkları için kayıptırlar.
Bazı evrimciler bugün bizim, insanın, kuyruksuz maymunlardan geldiğini asla söylemememiz, bunun yerine insanın ve kuyruksuz maymunların ortak bir atadan geldiklerini söylememiz konusunda ısrar etmektedirler. Bu gerçekten saçmalıktır ve insanın kökeninin, kuyruksuz bir maymundan geldiği fikrini halk tarafından daha kabul edilebilir hale gelmesi için ileri sürülmüştür. İnsanın, günümüz kuyruksuz maymunlarından gelmeyip günümüz kuyruksuz maymunlarının ve insanların ortak atasından geldiği görüşü evrimcilerin asıl görüşü olmasına rağmen, eğer içimizden biri bu kuramsal ortak atayı görmüş olsaydı, elbette ki onu bir kuyruksuz maymun olarak isimlendirirdik.
Bugün, Hominidae’de tek bir tür vardır: İnsan familyası –Homo sapiens– ya da günümüz insanı. Yaratılışçı bakış açısına göre insan, eşsiz yaratılmış bir canlı olarak daima diğer bütün canlılardan ayrı ve farklı olmuştur. Evrimsel bakış açısına göre, kendi soy oluş dalı milyonlarca yıl önce kuyruksuz maymunlardan ayrılmış insan, uzun bir evrim tarihine sahiptir. Kuyruksuz maymunlar ve insanlar arasındaki tüm ara form canlıları Hominidae’nin üyeleri olarak düşünülecektir ve bundan dolayı hominid olarak adlandırılmaktadır. Böylece evrimciler, günümüz insanı ve onun kuyruksuz maymunlarla olan son atası arasında pek çok ara form türünün bulunduğuna inanmaktadırlar ve Darwin’den bu yana süregelen bu varsayımsal geçiş formlarını bulma arayışları da yoğun olarak devam etmektedir.
Kendi türlerimizin kökenleri ile ilgili olarak ortaya atılan bazı ipuçlarının dışında başka hiçbir şey insanı daha fazla heyecanlandırmaz. Tanınmayan birden fazla paleoantropolog, insanın kökeniyle ilgili olduğuna inandığı bir canlının kalıntılarını bulduktan sonra ─ özellikle de bu bulgu Afrika veya Asya’nın tenha bölgelerinde bulunmuşsa ─ sansasyonel ve abartılı iddialarda bulunarak bir anda ün kazanmıştır. Göreceğimiz gibi bu tür iddiaların çoğu, yeni keşifler ve çalışmalar tarafından çürütülerek eninde sonunda karanlığa gömülmektedir. Birkaç durumda bu sansasyonel “bulgular”ın birer aldatmaca oldukları ortaya çıkmıştır.
Şekil 23’te, evrimcilerin, kuyruksuz maymunlar ile insanlar arası ara formlar olarak önerdikleri canlılar gösterilmiştir. Sözde evrimsel tarihin milyonlarca yıl sürdüğü inancına göre, önerilen ara formların sayısı oldukça azdır, üstelik bu ara formların bazıları da aile ağacından düşmeye başlamıştır.
Şekil 23. Evrimciler tarafından bugün önerilen tarihlerle, kuyruksuz maymunlardan insanlara doğru ilerleyen evrimsel çizgide varsayılan ara formlar.