5. BÖLÜM
FOSİL KAYDI ▬ BALIKLARDAN SÜRÜNGENLERE
Dört ayaklılar, amfibyumlar, sürüngenler ve memeliler içerisindeki tüm dört ayaklı hayvanları içine alırlar. Evrimciler, dört ayaklıların, bir balık atadan türediğini ve amfibyumların da ilk dört ayaklılar olduklarını var saymaktadırlar. Dört ayaklıların atası olan balığın hangi balık olduğu, evrimciler arasında çok tartışmalı bir konudur. Bu tartışmaların nedeni, paleontologların, balık ile amfibyum arası geçiş formu bulmada tamamıyla başarısız olmalarıdır. Eğer böyle geçiş formları bulunsaydı, atalar ile torunlar arasında bağlantı kurmak, oldukça basit bir iş olacaktı. Robert L Carroll, amfibyumların atasının, rhipidistian crossopterygian balığı (tanımlar için Sözlük bölümüne bakınız) olduğunu farz etmiştir,1 fakat şunu da açıkça kabul etmiştir: “Erken amfibyumlarla rhipidistian balıkları arası hiçbir ara seviye fosiline sahip değiliz....”2 Rosen, Forey, Gardiner ve Patterson bu önermeyi reddederek, amfibyumlara hayat verenin, bir akciğerli balık olduğu fikrini savunmaktadırlar.3 Thomas Gorr ve Traute Kleinschmidt ise bu iki önermeyi de reddedip, amfibyumların, bir coelacanth (silekant ya da koelakant) balığından evrimleştiğini savunurlar. Bu balığın soyunun, 80 milyon yıl önce tükendiğine inanılıyordu. Fakat, 1938’de, Afrika’nın doğu kıyılarında, balıkçılar tarafından, Latimeria cinsinin yaşayan bir örneği, bulununca, bunun doğru olmadığı anlaşıldı. Sonraları, denizden, yaklaşık 200 numune çıkarıldı. Bu iddianın dayandırıldığı gerçek şuydu: Latimeria’nın hemoglobin molekülü, akciğerli balık, köpekbalığı ve birçok teleost balığından çok, (teleostlar, bugünkü kemikli balıklardır), iri kurbağaların tetari türlerindeki Beta-hemoglobin zincirinin aminoasit dizilimine daha yakındı.4
Diğer yandan, Peter Forey, Latimeria’nın, amfibyumların atası olabileceği konusundaki iddiaları tümüyle reddederek şöyle der:
Coelacanth’ların, dört ayaklıların atalarına yakın oldukları gibi uzun süre kabul görmüş bir inancın var olması, Latimeria’nın da keşfiyle, balıkların amfibyumlara dönüşümü konusunda doğrudan bilgi elde edebilme ümitlerini arttırmıştır. Latimeria, dört ayaklılar ve coelacanth’ların, başka canlı gruplarına kıyasla, birbirleriyle çok daha yakın bir ilişki içinde oldukları kuramının dayandığı bir adla, “kayıp halka” ile tanıtıldı. Fakat, Latimeria’nın anatomik ve fizyolojik incelemeleri, bu ilişki kuramını kusurlu, yaşayan coelacanth’ların kayıp halka olarak isimlendirilmesini de, uygunsuz buldu.5
Bazıları, primatlar, sürüngenler ve dört ayaklılar gibi başlıca grupların kökenleri konusundaki çok sayıda literatürü eleştirel bir yaklaşımla okuduklarında, evrimcilerin bu tartışmalı ve çelişkili fikirleri karşısında kısa sürede bunalmakta ve yanılmaktadırlar. Her kuramcı, muhtemel atası olduğuna inanılan yaratığın seçiliminde en önemli olarak varsaydığı birçok özelliğin mukayesesine dayanan sözde mantıksal savını ileri sürer. Carroll, omurgalı paleontolojisi ve evrim konusunda yazdığı kitapta, rhipidistian balığının, amfibyumların atası olmasını sanki kaçınılmaz bir sonuçmuş gibi yazmaktadır. Yukarıda gördüğümüz gibi, başka evrimciler de akciğerli balık ve coelacanth’ların, en olası atalar olduklarında ısrar etmektedirler. New York Times’ta Malcolm Browne’nin yazdığı bir makalenin, Gorr ve Kleinschmidt tarafından yapılan eleştirisinde, onların, coelacanth’ların, amfibyumların atası olduğunu destekleyen fikirlerinden de söz edilmiştir.6 Browne ayrıca, Axel Meyer ve Allan C. Wilson tarafından yapılan ve coelacanthı olası bir ata olmaktan çıkaran en iyi aday olarak, akciğerli balıkları gösteren araştırmayı anlatır. Her ne kadar Carroll ve diğer evrimciler bunu gerçek gibi kabul etseler de, makalenin hiçbir yerinde, bir rhipidistian balığının amfibyumların atası olabilirliği konusunda tek bir kelime bile yoktur. Bu makalede Browne şöyle der:
Hangi balık grubunun ya da gruplarının karaya geçişte bir ilk olduğu ya da bunun evrimsel yolunun ne olabileceği konusunda hiç kimse emin olamaz....sudan karaya geçişin çok uzun zaman önce olması ve fosil kaydından çıkarılan çeşitli soy ağaçlarının böylesine karışık olması, evrimcilerin, bunları kesin olarak çözümleyemeyecekleri olasılığını kabul etmelerini sağlamıştır.
Yani, Carroll rhipidistian balığını, Gorr ve Kleinschmidt coelacanthı, Rosen ve diğerleri de akciğerli balığı seçmekte (onaylamakta) ve her biri de diğer önermelerin niçin geçersiz olduğu konusunda nedenler ortaya koymaktadırlar. Bu karışıklık ve tartışmanın kaynağı nedir? Her zaman olduğu gibi, geçiş formların yokluğu yüzündendir! Sadece birkaç geçiş form, amfibyumların atasını ve evrim yolunu gün ışığına çıkaracaktır. Geçiş formlar yokken, tüm önermeler sadece birer senaryo ve boş konuşma oluyor.
Zamanının tümünü suda geçiren bir balığın, zamanının büyük oranını ya da çoğunluğunu karada geçiren bir canlıya dönüşümü, ataların amfibyumlara dönüşümü sırasında geçirdikleri morfolojik ya da biçimsel anlamda basit değişimlerden çok daha fazlasını gerektirecektir. Carroll, bir balığın, bir amfibyum olabilmesi için birkaç önemli değişim geçirmesi gerektiğini söyler.7 Bunların arasında şunlar vardır:
Su Dengesi: İlkel amfibyum, sudan karaya hareket ettiğinde, ağız, akciğer ve vücut yüzeyi bölgesinde gerçekleşecek ciddi bir su kaybına uğrayacaktır. Bu kayıpları en aza indirgeyebilmek için, bu yapı ve organlarda önemli değişimler gerçekleşmek zorundadır.
Duyu Organları: Bir balık, ya da var sayılan ilkel amfibyum, su ile hava arasındaki fiziksel ve kimyasal farklara uyum sağlayacak, duyusal bir yeniden yapılanma geçirmeden nasıl hayatta kalabilecektir? Unutmayın ki, her duyu organı başlangıçtan itibaren doğru çalışmalı, her değişim doğru bir sırada gerçekleşmeli, diğer tüm değişimlerle uyumlu olmalı ve tüm bunlar, genlerin, tesadüfi, rastlantısal yollarla değişmesi, yani mutasyon geçirmesiyle gerçekleşmelidir. Yeni gelişmeye başlayan amfibyumun ya da kısmi amfibyumun karada yaşamasını mümkün hale getirecek daha başka pek çok önemli fiziksel değişim gerçekleşmiş olmalıdır.
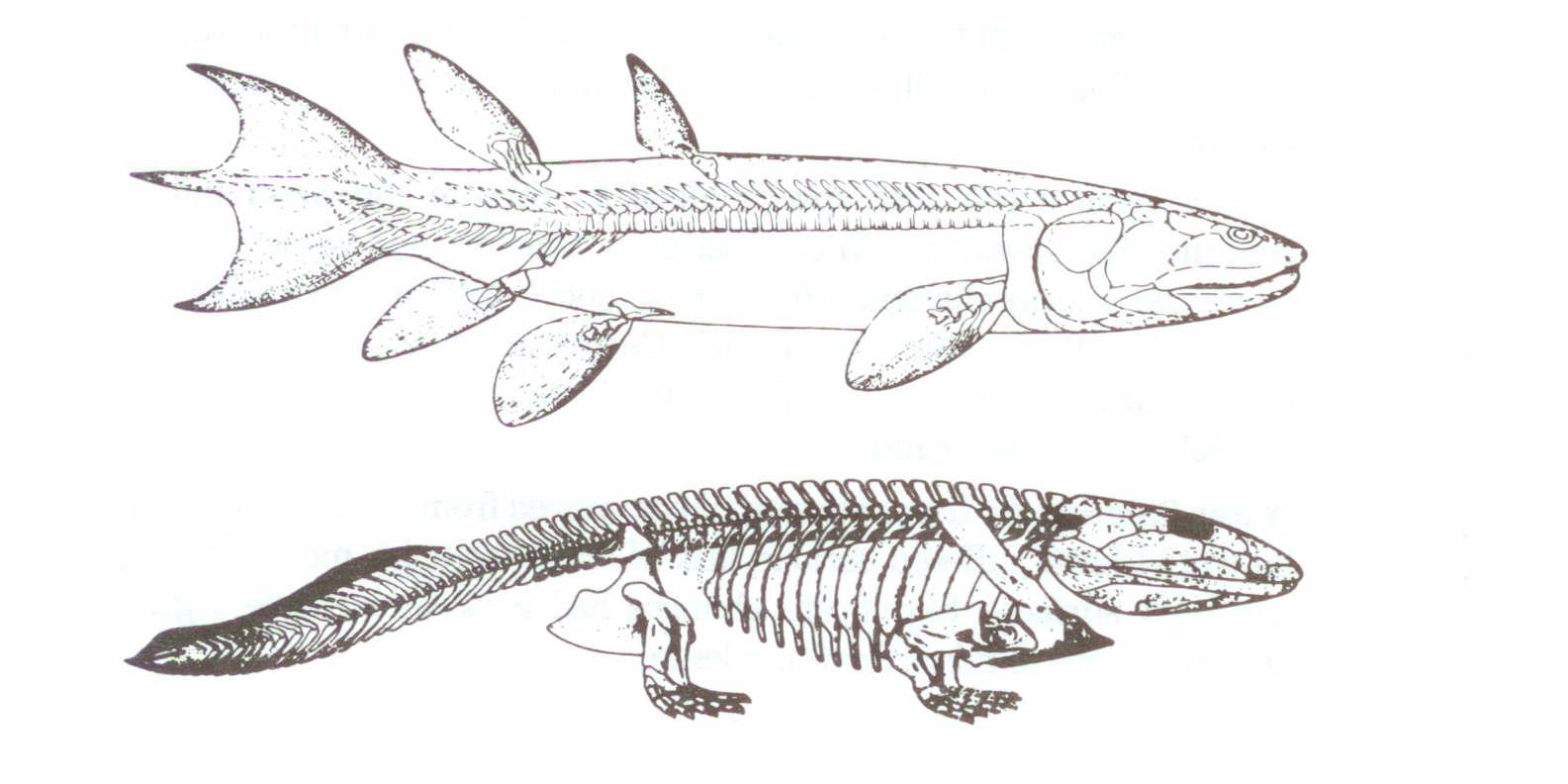
Bir balığın, bir amfibyuma dönüşümü için, çok önemli morfolojik değişimler olması gerekmektedir. Balık yüzgeçleri, suda hareket, yön ve dengeyi sağlayıcı yapıda tasarlanmıştır. Yüzgeçlerin, su tarafından sağlanan kaldırma gücünün olmadığı havada, balığın vücudunu desteklemesi olası değildir. Bir balığın pelvis kemiği, omurgayla hiçbir bağlantı olmaksızın, gevşekçe kasların içine gömülmüştür. Omurgaya bağlantı gerekli değildir.Yüzgeçler, vücut ağırlığını desteklemezler. En eski amfibyumlardan biri olduğu tahmin edilen Ichthyostega’da, (Şekil 3), görünüşe göre, pelvis kemiği, oldukça geniştir ve omurgaya da sıkıca bağlanmıştır. Bu, karada hareket için geçerli olan anatomi biçimidir. Bir su altı aracından alınan derin deniz fotoğrafları, bazı evrimcilerin söyledikleri şeylerle çelişkili olarak, coelacanth Latimeria’nın, suda kaldırma gücünü kullanabileceği gerçeğine rağmen, okyanus dibinde tek bir adım dahi atmadığını ortaya çıkarmıştır. Burada dikkat edilmesi gereken bir önemli nokta da, bir balığın bir amfibyuma kuramsal olarak evrimleşmesi esnasında, bir nedenden dolayı, bu dönüşümün, sırt ve anüsle ilgili iki yüzgeci de bırakmasıdır. Evrimciler, balık benzeri kuyruk yüzgecinin varlığının, Ichthyostega’nın, rhipidistian balığı soyundan geldiğini gösteren bir kanıt olduğuna işaret ederler. Fakat, Forey’in de vurguladığı gibi, balık benzeri kuyruklar, (Gnathostomata) çeneli omurgalı gruplarının genel karakteristikleri olduğu için, bu kanıt geçersizdir.
Şekil 3: Bir ichthyostegidamfibyumunun yapılanışı ve onun sözde atası olan crossopterygian. Romer’in Vertebrate Paleontology kitabından, Chicago Üniversitesi
Yayınları’nın izniyle.
Carroll şöyle der:
Rhipidistian’lar ve amfibyumlar arasında en göze çarpan farklılıklar, etraf (appendicular) iskeleti üzerindedir....Erken amfibyumlarda, kol bağlantıları ve kolun merkezden uzak (distal) parçaları önemli biçimde şekil değiştirmiştir.8
Carroll, rhipidistian’ların kalın, loplu-çift yüzgeçlerinin işlevlerinin, amfibyumların ayak ve bacak işlevlerinden çok daha farklı olduğunu belirtti. Rhipidistian balığında kol kemiği (ön yüzgeç çifti), ve kalça kemiği (arka yüzgeç çifti) vücuda, yanal ve dönel hareketleri kısıtlayacak derecede yakın bulunmaktaydı. Yüzgeçler, kıç tarafına yönelmişlerdi. Ancak, “ilkel” dört ayaklılarda, kol ve bacak gibi üyeler, hareket esnasında öne ve gövdeden yanlara uzuyordu. Böylece, amfibyumların hareket biçimi, “en eski” ya da en “ilkel” amfibyumlarda bile herhangi bir balığınkinden çok farklıydı.
Fosilbilimciler, crossopterygian balığının göğüs yüzgecinde bulunan kemiklerin, amfibyumların ön kollarında bulunan pazı, önkol ve dirsek kemiklerine benzer olduğu gerçeğini işaret etmişlerdir. Fakat bu benzerlik, balıkların göğüs ve kalça yüzgeçlerinin, amfibyumların ön ve arka bacak üyelerine benzerliğine kadar erişemeden son bulmaktadır. Bu balıkların herhangi birinde bile, amfibyumların el ve ayaklarına eşit sayılabilecek ya da benzetilebilecek hiçbir şey yoktur.
Carroll şöyle der:
Merkezden daha uzakta, yüzgeçleri destekleyecek hiçbir geniş iç iskeletsel yapı yoktur; ve insanlar, dört ayaklıların geliştirdiği el tarağı ve parmak kemikleri gibi yapıların tümüyle olmasa da tüme çok yakın yeni yapılanmalar olduğunu kabul etmelidirler.9
Ahlberg ve Milner şöyle der:
Dört ayaklıların kol ve bacak üyelerinin bağlanma noktalarına yakın üst üyeler, sarcoptergian balıklarınınkine çok benzerler. Ancak, Şekil 5’te de görüldüğü gibi, süreç, “yön değiştirerek” dirsek kemiğinin uzantıları olan parmakları üretir. Loplu yüzgeçli balıklarda gerçekleşmiş buna benzer hiçbir süreç bilinmemektedir. Görünüşe göre, eller ve ayaklar, çift uzantıların bağlanma noktalarından uzak parçalarını etkileyen geliştirici bir değişim sonucu üretilmiş “yeni” yapılardır.10
(Ahlberg ve Milner tarafından yazılan makale, bir eleştiri makalesi olup, bu yazılışta evrimcilerin, dört ayaklıların kökeni ve erken çeşitliliği konusundaki en yeni fikirleri anlatır.)
Daha yüksek dört ayaklı grupların kökenleri11 konusunda son yıllarda yazılan bir kitabın, Hans-Peter Schultze’ye ait bölümü, uygun bir şekilde “Dört Ayaklıların Kökeni Konusundaki Tartışmalı Hipotezler” olarak adlandırılmıştır. Schultze, son söz olarak şöyle der:
Aynı özelliklerin analizlerinden türetilen dört farklı filojeni (soyoluş) sırası tartışılmıştır. Her yazar, karakterleri, kendisinin tercih ettiği dört ayaklı ilişkisinin tarafını tutarak yorumlamıştır. Jarvik, gerçek dört ayaklılar ve kuyruklu amfibyumlar arası bir farkı vurgularken, aynı zamanda, kuyruklu amfibyumlar ile delik pullu (porolepiform) balıkları arası benzerliklere de dikkat çekmiştir. Rosen ve diğerler kemik pullu (osteolepiform) balıkları ve dört ayaklılar arası benzerliklerin önemini küçümsedikten sonra, akciğerli balıkların özelliklerini dört ayaklılar biçiminde yorumlamıştır. Chang, akciğerli balıkların da içinde bulunduğu tüm sarcopterygian balıklarına huni şeklindeki burun boşluğunun (choana) dahil olmadığı fikrini savunmuştur. Zaten, Chang’a göre, dört ayaklıların tarihi, diğer tüm sarcopterygian’larınkinden farklıdır. Vorobyeva ve Schultze, osteolepiform’lar ile panderichthyid’lerin iç damak başlangıçlarının gerçek bir choana olduğunu ve dört ayaklılara kıyasla, yüzgeçlerin iç yapılarının sadece osteolepiform’larda böyle olduğunu kabul ettiler.... bütün akrabalık meseleleri, benzerliklerin belirlenebilmesi için özelliklerin benzerliklerinin ya da benzemezliklerinin değerlendirilmesine dönüşür.... Rosen ve diğerleri (1981) akrabalık analizlerinin, öncelikle, günümüzde hâlâ var olan, analizlerde ikinci derece fosil olarak tanınan formlara dayandırılması gerektiğini tartışmıştır. Fosil kaydındaki kusur, herkes tarafından kabul edilmektedir. Sonuç olarak fosiller, filojenetik düzen çözümüne hiçbir katkıda bulunmamaktadırlar.12
Schultze ve diğerleri, her düzenin ortaya çıkardığı çelişki ve güçlükleri açıklayabilmek için, her yazarın, çok dolaylı istatistikler yapması gerektiğini belirtmişlerdir. Yukarıda da belirtildiği gibi, Schultze şöyle der: “bütün akrabalık meseleleri, özelliklerin benzerliklerinin ya da benzemezliklerinin değerlendirilmesine dönüşür....” Fakat pek çok örnekte, evrimcilere göre, benzerlikler, atasaltorun ilişkileri konusunda hiçbir şeyi ispat etmemektedir. Çünkü, farklı hayvanlarda benzer yapılar bu hayvanların genlerinin ortak bir atadan gelerek değil, bu özellikleri birbirinden bağımsız olarak kazanmışlardır. Evrimciler, bir özelliğin, yakın akraba türlerde bağımsız olarak evrimleştiğini farz ettiklerinde, Paralel Evrim diye isimlendirdikleri bir yola başvurmaktadırlar. Evrimcilere göre, bu, evrimde yaygın bir var oluştur. Evrimcilere göre, evrimdeki bir diğer yaygın fenomen de Yakınsak Evrim (convergent evolution) diye isimlendirilir. Buna göre, uzak akraba türlerin bağımsız anlamda benzer yapılar ve özellikler geliştirdiği varsayılmıştır. Sıkça sözü edilen bir örnek de, omurgalıların ve omurgasız kafadanbacaklıların (mürekkepbalıkları) gözleridir. Bu iki canlının gözleri, birbirine çok benzer. Fakat bu iki tür canlının varsayılan ortak ataları, hem kafadanbacaklıların hem de omurgalıların gözleri gelişmeden yüz milyonlarca yıl önce var olmuş olmalıdır. Futuyma şöyle der:
Yakınsak evrim, pek çok örnekte öyle benzer özelliklerle sonuçlanmaktadır ki, bizler, eğer bu özelliklerin akraba olmayan gruplarda bağımsız olarak evrimleştiğini bilmiyor olsaydık, aynı genetik temele sahip olup olmadıklarını söylemekte de güçlük çekecektik.13
Böylece, farklı hayvanlardaki benzerlikler, ortak bir atadan evrimleşmenin kanıtı olabilirler de olmayabilirler de. Çünkü, bu özellikler, paralel ya da yakınsak evrim yoluyla bağımsız bir biçimde ortaya çıkmış olabilirler. Evrim kuramında, evrimsel tasarımı daha karmaşık bir hale getiren, fakat bu tasarımların korunmasında da başvurulan bir diğer yaygın özellik de tersine dönüş olarak isimlendirilen özelliktir. Bu tersine dönüş fikri, bir neslin evrim tasarımındaki bir yapı kaybolduğu ve daha sonra torun varsayılan türlerde yeniden ortaya çıktığı durumlarda evrimciler tarafından ortaya atılmıştır. Uzun süre, evrimde, tersine dönüşün olamadığına inanıldı. Karmaşık bir yapının kökeni ile ilgili çok sayıda genetik değişimin olması gerektiği ve bu değişimlerin karmaşık bir sıra halinde kazanılması gerektiği iddia edildi. Bir sonraki evrimsel değişimle yok edilen yapının yeniden geri kazanılması ihtimali aslında hiç yoktur. Gerçekte, bu prensibin öyle iyi saptandığı düşünülmüştür ki, prensip, “kanun” statüsüne yükselmiş ve Dollo Kanunu olarak isimlendirilmiştir. Şimdi, Dollo’nun ileri sürdüğü kanunu, modern evrim tasarımında sık sık başvurulan tersine dönüşlerde olması gerektiği gibi, bir çöp kutusunun içine fırlatılıp atılmıştır. Örneğin, Schultze şöyle der:
Yukarıda açıklanan türdeki tersine dönüşler, pek çok kafatası ve daha alttaki özelliğin açıklanmasında da gereklidir. Kafatası çatı kemiklerinde, actinopterygian'lardaki parietal ve postparietal’lerin açıklanmasında iki, dört ayaklılar ve sarcopterygian'ların ortak atalarındaki alın kemikleri, parietal ve postparietal’lerin açıklanmasında üç, Diabolepis, porolepiform’lar ve actinistian’lardaki parietal ve postparietal’lerin açıklanmasında da iki tersine dönüş gerekmektedir. Filojenetik sırada, ikiden bire ve sonra tekrar ikiye yükselen squamosal ve dış geniz açıklığı ile yok olan, var olan ve sonra tekrar yok olan omur ve choana ifade edilmektedir.... Sarcopterygian balıkları ve dört ayaklılar arasındaki bir kardeş ilişkisi, ortak atanın, balık benzeri torunlardan türemiş bir dört ayaklıya çok daha fazla benzediğini ortaya çıkaracaktır. Böyle bir düzen, olası gibi görünmeyen sayısız karakterin tersine dönüşünü gerektirecektir.14
Daha önceden de belirtildiği gibi, Schultze şöyle demiştir: “her yazar, karakterleri, kendisinin tercih ettiği dört ayaklı ilişkisinin tarafını tutarak yorumlamıştır.” Diğer bir deyişle, her araştırmacı, en başta kendisinin tercih ettiği bir varsayımsal evrim modelinde karar kılar ve sonra düzenin gerektirebileceği tersine dönüşler, yakınsak evrim ya da paralel evrimi kullanarak, kanıtları uygun biçimde yorumlar. Bu kuramlar, öylesine istenilen şeklin verilebildiği kuramlardır ki, yanlış olsalar bile doğru ya da yanlış oldukları ispat edilemez ve bu nedenle, bilimsel kuram olarak bile isimlendirilemez. Ahlberg ve Milner, Rosen ve diğerlerinin, dört ayaklıların kökeni konusunda15 yayınlanmış olan fikirleriyle ilgili olarak şöyle dediler:
Onların, dört ayaklıların, osteolepiform’lardan evrimleşmesinin, kalıntılarına bakarak geçmişin anlaşılabileceğini gösteren evrim senaryoları, kesinlikle doğruluğu ya da yanlışlığı ispat edilemeyen “hikayeler” olup bilim değildir.16
Kemik pullular (Osteolepiform), çift yüzgeçli olan Crossopterygii alt sınıfının Rhipidistia takımı içinde yer alan balıklardı ve evrimcilerin çoğunluğu tarafından bunların, amfibyumların atası olduklarına inanılmaktadır.
Pennsylvania’nın Catskill Oluşumu’nun Üst Devoniyen kayalarında bulunan bir amfibyum fosili bulgusu, en eski amfibyumlar olduklarına inanılan bu canlıların, hiçbir geçiş formu kanıtı olmaksızın tamamıyla gelişmiş oldukları delillerini güçlendirmiştir. Bu kayalar, evrimcilere göre, günümüzden yaklaşık 363 ile 365 milyon yıl öncesine ait Orta ve Üst Famennian oluşumlarındadır. Bu amfibyum hakkındaki rapor şöyleydi: Hynerpeton bassetti olarak isimlendirildi, Science’in Daeschler, Shubin, Thomson ve Amaral17 tarafından yapılan 29 Temmuz 1994 basımında yer aldı. Onlar şöyle dediler:
Omuz kemerinden türemiş özellikler, uzantılı mekanizma desteğinin ve itici kuvvetinin, dört ayaklılar tarihinin en erken dönemlerinde bile çok iyi gelişmiş olduğunu göstermektedir.
Fosilin bulunduğu katmanın açıklanmasında bu yazarlar şöyle dediler:
Bu katman durumu, H. bassetti’nin Orta–Üst Famennian yaşında olduğunu, (yaklaşık 363-365 milyon yıl), yani, en üst Famennian yaşında olan Tulerpeton, Acanthostega ve Ichthyostega’dan daha yaşlı olduğunu gösterdi.... Ayrıca, H. bassetti’nin omuz kemeri morfolojisi, onun, ön kollarını kaldırma, tepki verme ve güçlü bir şekilde uzatma yeteneğine sahip olduğunu gösterdi.
Gördüğümüz şey, tamamıyla gelişmiş vücut üyelerine ve gelişmiş diğer özelliklere sahip, %100 bir amfibyum olan ve önceden, en eski amfibyumlar olduğu düşünülen canlılardan daha yaşlı bir canlıdır. Ayrıca bu, evrim kuramının dayandırıldığı tahminleri yalanlamakta, fakat, yaratılış için, güçlü, ek kanıtlar sağlamaktadır.
Öncelikle belirtmek istiyoruz ki, amfibyumları ve böylece “ilk” dört ayaklıları bir balık atadan türetmeye çalışan tüm projeler, bilimsel kuram olarak nitelendirilemeyen senaryolardan başka bir şey değildirler. Üstelik, balıklarla amfibyumlar arasında tasarlanan evrimsel yol ne olursa olsun, tüm balıklar ile amfibyumlar arasında büyük bir devamsızlık vardır. Amfibyumların en olası ataları olarak sözü edilen balıkların tümü, zamanlarının tümünü, su içinde geçirmeye ihtiyaç duyan, %100 balık olan balıklardı. Soylarından geldiği düşünülen amfibyumların tümü ise, temel amfibyum kol ve bacak üyelerine sahip, %100 amfibyum olan amfibyumlardı. Hiç kimse, yarı ayak yarı yüzgeç üyeleriyle ilgili tek bir geçiş formu bulmakta başarılı olamadı. Bu tür şeyler bulunmuş olsaydı, amfibyumların atası olma konusunda rekabet halinde olan adaylardan hangisinin gerçek ata olduğu hemen belirlenmiş olacaktı. Fakat tartışmalar dinmemektedir. Geçiş formları bulunamamıştır; çünkü, asla var olmamışlardır. Yeryüzünde, canlı organizmalar evrimleşmemiştir.
Amfibyumların Büyük Çeşitliliği
Günümüz amfibyumları, 4.000’den fazla türü ile, büyük bir çeşitlilik göstermektedir. Bu 4.000 tür, üç takımı kapsamaktadır: Urodela (semenderler ve su kelerleri); Gymnophiona (kertenkelemsigiller, caecilian ve apodanlar olarak da isimlendirilen hiç bacak izlerine sahip olmayan, solucan benzeri amfibyumlar); ve Anura (kurbağalar ve kara kurbağalar). Bunlar, fosil kaydında ilk ortaya çıktıklarında, tıpkı bugün oldukları gibi, modern bir görünüş ve yapısal olarak büyük bir çeşitlilik içeriyorlardı. Carroll şöyle der:
Kurbağalar ve semenderler, fosil kaydında ilk ortaya çıktıklarında, esasen modern bir iskelet anatomisi gösteriyorlardı. Gymnophiona’nın, (caecilian), açıklanmış olan fosilleşmiş canlıları, günümüz cinslerine çok benzeyen, Üst Kretase ve Paleosen katmanlarında ayrık halde bulunan omurgalarıyla tanınır.18
Bu modern amfibyum takımları, genellikle Lissamphibia alt sınıfı içinde yer almışlardır; çünkü, birkaç benzerlik paylaşmaktadırlar. Ancak, pek çok paleontolog, böylesine çeşitlilik içeren canlıları birlikte gruplandırma konusunda şüphelidirler. Carroll şöyle diyor:
Benzerliklerine rağmen, kurbağalar, semenderler ve kertenkelemsigiller (caecilian’lar), iskelet yapısı, yaşam biçimi, şimdiki ve bilinen tüm fosil kaydı bakımından birbirlerinden çok farklıdırlar.... biz, günümüz takımının her üçünde de ortak olup özelleşmiş niteliklere sahip olan herhangi bir olası atanın hiçbir fosil kanıtına rastlamadık.... Kurbağalar, semenderler ve kertenkelemsilerin yakın bir ortak atadan evrimleşmesi konusundaki fosil kanıtı yokluğunda, günümüz takımlarının her birinin, bir grup bağımsız Paleozoik amfibyumundan evrimleşmesi olasılığını dikkate almalıyız.19
Benzer şekilde, Colbert ve Morales şöyle derler:
Bu benzerliklere rağmen, tek bir ortak atadan beklenecek bir özellik olan, karakteristikleri bir arada toplama özelliğini taşıyan herhangi bir Paleozoik amfibyumun kanıtı yoktur. En eski kurbağalar, semenderler ve kertenkelemsiler, günümüzde yaşayan torunlarına çok benzemektedirler.20
Carroll,21 Colbert ve Morales,22 kurbağa benzeri bir Triadobatrachus’tan bahsederken, günümüz kurbağaları ile diğerleri, yani, eski kabul edilen amfibyumlar arasında olası bir halka olduğunu ileri sürerler. Carroll, Triadobatrachus’un, Paleozoik amfibyumlarına olan en yakın benzerliğinin, dissorophid temnospondyl olan Doleserpeton’da, olduğunu söyler. Ancak, tüm kurbağalar ve gerçekte, tüm günümüz amfibyumları, temnospondiller ve en yaşlı kabul edilen tüm amfibyumlar arasında temel bir fark vardır. En yaşlı oldukları iddia edilen amfibyumlar, her bir omuru oluşturabilmek için, birbirine bağlanmış, pek çok farklı parçadan oluşan, kemer biçimli, karmaşık bir omura sahiptiler. Ayrıca, yine amfibyumların olası atası olarak önerilen çeşitli balık türleri de buna benzer karmaşık biçimli bir omura sahiptiler ve bu, en yaşlı kabul edilen amfibyumlarla balıklar arası bir halka için anlamlı karakteristiklerden biridir. Günümüz amfibyumlarının tümünün çok farklı türden, makara benzeri, kemikli bir silindire benzeyen omurları vardır. Eğer Doleserpeton gibi bir canlı, kurbağaların ya da diğer günümüz amfibyumların herhangi birinin atası olsaydı, Doleserpeton’un karmaşık, kemer biçimindeki omuru, basit makara biçimli omura dereceli olarak evrimleşmiş olmalıydı. Fosil kaydında, bu konuda, kesinlikle hiç kanıt yoktur.
Makara biçimli omura sahip amfibyumlar, lepospondyl olarak isimlendirilmektedir. Mississippian ve Pennsilvaniyen kayaları arasında makara biçimli omura sahip amfibyumların üç farklı takımı bulundu. Bu takımlar, Aistopoda isimli, en fazla 230 omurdan oluşan uzun bir vücuda sahip yılan benzeri canlıları içeriyorlardı. Bu canlılarda ne bacak ne de bacak kemeri bulunmuyordu. Carroll şöyle der:
Bu formların, yüksek derecede özelleşmiş kafatasları vardı. Bu, her familyanın, hem herhangi bir labyrinthodont’tan ya da rhipidistian’dan farklı hem de birbirinden farklı, uzun bir tarihi olduğunu gösteriyor.23
Aistopod’lar hakkında konuşan Romer şöyle der:
Bazılarımız, böylesine özelleşmiş formların, daha yaşlı amfibyum grupları arasındaki bir geç gelişimi temsil edeceğini ümit edebilir; fakat henüz tanımlanmayan yeni bulgular, aistopod’ların, Mississippian’in erken dönemlerinde geliştiğini ve bu yüzden bilinen tüm amfibyumların en yaşlıları arasında olduklarını kanıtlamaktadır.24
Gerçekte, Alt Mississippian kayalarından çıkarılan aistopod’lar, şimdi çok iyi tanımlanmış oldular. Bu amfibyumlar, fosil kaydında, herhangi bir eski amfibyumdan farklı olarak, tamamıyla gelişmiş biçimde aniden ortaya çıkmışlardır. Bunlar, alt (erken) Mississippian kayalarında, tamamen gelişmiş biçimde ortaya çıktıklarına göre, bunun anlamı şuydu: Eğer bunlar evrimleşmişlerse, Devoniyen dönemde evrimleşmiş olmalıydılar. Bununla beraber, en erken ya da en eski olarak isimlendirilen evrimsel balık soyu olan amfibyumlar da Devoniyen’de bulunmuştur. Eğer evrim senaryoları doğru ise, Ichthyostega gibi amfibyumların yakın ataları, balık atalarından bacakları evrimleştirmekle meşgulken, balık ile aistopod arası ara formlar da bacak gibi üye izlerini yok etmekle meşguldüler. Bazı amfibyumlarda bacak ve ayakları bir evrimsel doğuşa yönlendirirken, aynı zamanda da diğer amfibyumların bacaksız oluşmalarını nasıl bir seçim baskısı sağlayacaktı?
Üstelik, eğer, Ichthyostega gibi amfibyumlarda bulunan karmaşık, kemer tipi omur, rhipidistian balığındaki kemer tipi omura benziyorsa ve ikisi arasında bir bağlantı oluşturuyorsa, o zaman, basit, makara biçimli omura sahip olan lepospondyl amfibyumları nereden evrimleşmişlerdir? Bir balığın, tamamıyla basit, makara biçimli, 230 omurdan oluşan, yılan benzeri bir vücuda sahip, bacak olmayan bir aistopoda evrimleşmesini gösteren geçiş formları nerede? Bunlar, evrim kuramının bütünüyle iflasını ortaya koyan gerçeklerdir.
Aistopoda ile birlikte ortaya çıkan diğer iki lepospondyl amfibyum takımı, Nectridea ve Microsauria’dır. Nectridean’ların, çok küçük üyeleri olan uzun vücutları vardı. Yüzme organı olarak kullandıkları, yana doğru basık kuyrukları olan bu canlılar, görünüşte, çoğunlukla suda yaşarlardı. Microsaurian’lar, üyeleri oldukça küçük olsa bile, daha normal bir vücut yapısına sahiptiler. “Küçük sürüngenler” anlamına gelen Microsauria ismi, uygun bir isim değildir. Çünkü, Romer’in de söylediği gibi, sürüngenlere olan bu benzeyiş, sadece bir paralellikten ibarettir. Microsauria’lar, çeşitli vücut bölümleriyle, lepospondyl’lerin en çeşitlileridir. Aistopoda, Nectridea ve Microsauria, Mississippian, Pennsilvaniyen ve Permiyen kayalarında bulundular ve lepospondyl’lerin Paleozoik (Birinci Zaman) grubu olarak nitelendirildiler.
Evrimsel zaman ölçeğinde, Paleozoik lepospondyl’lerin ve Lissamphibia Altsınıfının günümüz amfibyumları arasında çok büyük bir boşluk vardır. Romer, Lissamphibia hakkında konuşurken şöyle demiştir:
Onlar ve Paleozoik grubu arasında fosil kaydıyla birleştirilemeyen geniş bir evrimsel boşluk vardır.25
Carroll şöyle yazar:
Alt Permiyen’den sonraki herhangi bir lepospondyl fosili bulamadık. Kurbağa ve semenderlerin Jura’da ortaya çıkışlarına kadarki küçük amfibyum kayıtlarında şaşırtıcı bir boşluk vardır.26
Kesinlikle açık olan şey, amfibyumların fosil kaydının, dört ayaklıların ve özellikle amfibyumların bir balık atadan evrimleştiği fikrini desteklememesidir. Rhipidistian crossopterygian’lar ya da diğer benzer balıklar ile, ichthyostegid’ler ya da diğer “en erken” amfibyumlar arasında aşırı derecede büyük bir boşluk vardır; gerçekte, tüm Paleozoik amfibyum takımlarının, takıma ait karakteristiklerin büyük çeşitliliği ile aniden ortaya çıkışı, ilk numunelerle bütünleşmektedir. Ve bu Paleozoik takımları ile üç canlı takımı arasında geçiş formu yokluğu, bu formların, evrimsel bir geçiş yoluyla ortaya çıktıklarını, kesinlikle inanılmaz hale getirmektedir. Yani, gerçekler, yaratılış modelinin dayandırıldığı tahminlerle tam bir uyum içerisindedir.
SürüngenlerinKökeni
Sürüngenlerin kökeni konusunda da, amfibyumlarda bulunan aynı durumla karşılaşıyoruz: Tahmin edilen evrimsel bağlantıları sağlayan ve temel biçimleri yönlendiren evrimsel yolları açığa çıkaran hiçbir geçiş formu olmaksızın, temel türlerin aniden ortaya çıkışı.
Sürüngenler, Reptilia sınıfı içerisinde; sürüngenler, kuşlar ve memeliler ise Amniota grubunda yer alırlar. Bu amniyonlular, amfibyumlardan farklı olarak, yumurtalarını su ve nemli ortamlardan uzakta bırakabilirler. Çünkü onların yumurtaları, embriyon etrafında zarla donatılmıştır. Amniotik bir sürüngen yumurtası, bir amfibyum yumurtasından çok daha karmaşıktır. Sürüngen yumurtaları, embriyonun gelişimi için, içinde bir kese bulunduran amniyon adlı bir zar içerirler. Alantoyis (Allantois), artık ürünlerin toplandığı bir depo görevi görür ve içinde gelişmekte olan embriyona yiyecek sağlayan bir yumurta sarısı kesesi vardır. Yumurta, içeriğini koruyacak derecede güçlü bir kabukla tümüyle sarılmıştır. Fakat aynı zamanda, bu kabuk, oksijenin içeri alınıp, karbondioksitin dışarı salınması gibi çevresel gaz değişimlerine olanak sağlayacak derecede gözenekli bir yapıya sahiptir. Evrimciler, bu tümüyle karmaşık sistemin, bir şekilde, adım adım, amfibyumun genetik sisteminde gerçekleşen bir dizi tesadüf ve yönü belli olmayan değişimler ile evrimleştiğini hayal etmiş olmalılar. Bu değişimlerle birlikte, ilkel sürüngenin üretim sisteminde gerekli olan değişimler de meydana gelmeli, yine atasal amfibyumun genetik sisteminde tamamen tesadüflere dayalı değişimler sonucu üretilmiş olmalıdır. Dönüşüm süresince, her ara seviye, kendisinden önceki seviyenin yerini alabilmesi için, kendini önceki seviyeden üstün kılan yararlar sağlamalı ve tümüyle işlevsel olmalıdır. Amniotik boşluk ve amniyon gibi hayati yapıların birçoğu, sürüngendeki bir hiçten üretilmiş olmalıdır. Çünkü amfibyumlarda, sürüngenlerin türeyebileceği herhangi bir benzer (homolog) yapı bulunmamaktadır. Olasılıkla, hiçbir evrimci, amniotik yumurtanın evrimsel kökenini sağlayan evrimsel yolları ve ara seviyeleri hayal etmeye bile kalkışmaz.
Evrimciler, belirli amfibyum ve sürüngenlerde, sürüngen ile amfibyum arası bir bağ oluşturan birçok karakteristiğin var olduğuna inanırlar. Örneğin, Colbert ve Morales şöyle der:
Seymouria’da gözlemlenen amfibyum ve sürüngen karakteristiği karışımı, omurgalıların evrimi sırasında iki sınıf arasında gerçekleşmiş olan kademeli dönüşümün göstergesidir.27
Seymouria’yı, sürüngen–amfibyum arası bir olası ara seviye gibi göstermeye yönelik herhangi bir girişim, derhal ciddi çelişkilerle karşı karşıya kalmaktadır. Bilinen en eski sürüngenler oldukları iddia edilen canlılar, Hylonomus ve Paleothyris, sırasıyla, Alt Pennsilvaniyen (evrimsel zaman ölçeğine göre günümüzden yaklaşık 315-330 milyon yıl önce) ve Orta Pennsilvaniyen (yaklaşık 310-315 milyon yıl önce) kayaları arasında bulunmuşlardır. Alt Permiyen kayaları arasında bulunan Seymouria fosillerinin yaşı 280 milyon yıl olarak belirlenmiştir. Bunun anlamı, bu fosillerin, sürüngenlerin atası olabilmek için en az 25-30 milyon yıl daha genç olmalarıydı.
Üstelik, Seymouria’yı sürüngenlerle bağdaştırma girişimlerinde başka ciddi problemler de vardır. Romer, şu soruyu sormaktadır: “Seymouria, yumurtalarını kurbağalar gibi suya mı bıraktı yoksa amniyonlular gibi yumurtalarını kabukla kaplayıp karaya mı bıraktı?” Romer, bu sorunun cevabının, Seymouriamorpha takımının içindeki çok sayılı canlıların incelenmesiyle bulunabileceğini söyleyerek, Kotlassia ile ilgili olarak konuşmasını şöyle sürdürür:
....yassı kafataslarıyla, iskeletleri nispeten zayıf, tipik amfibyumlarınkine benzeyen, kalıcı su yerleşiminin varlığına doğru, bir dizi ters yöndeki değişimler olarak kendini gösterir. Yine, Moravya’nın erken Permiyen döneminde keşfedilen Discosauriscus’un solungaç taşıyan larvalarının, rhachitome’lerden “branchiosaur” larvalarına benzer ve seymouriamorph’ların üretken özelliklerinin, kesinlikle amfibyum olduklarını gösterir.28
Diadectes’lerin amfibyumlar ile sürüngenleri ayıran çizgiye çok yakın canlılar olduklarından sıklıkla bahsedilmektedir. Oysa, bu canlı, aynı zamanda, bu çizgide var olma olasılığı çok düşük bir canlıdır. Diadectes, evrimsel zaman ölçeğine göre, sürüngenlerin atası olabil-mesi için yaklaşık 30 milyon yıl daha önce yaşamış olması gerekirdi. Üstelik bu canlı, burundan kuyruk birleşimine kadar yaklaşık on cm uzunluğundaki “ilkel” sürüngenlerin aksine, üç metre uzunluğunda, büyük, hantal bir hayvandı.
Seymouria ve Diadectes’in birçok karakteristiğinin, Hylonomus ve Paleothyris ve diğer “ilk” sürüngenlerdeki benzerlikleri açığa çıkardığı söylenmesine rağmen, amfibyumlar ve sürüngenler arasında, hiçbir bağlantısı olmayan bir boşluk vardır. Sürüngenleri, atasal amfibyumlar olarak isimlendirilen canlılara bağlayan hiçbir geçiş formu yoktur ve sürüngenlerin, başlangıçta da sürüngen olduklarını ortaya çıkartan birkaç örnek aşağıda verilmiştir. Omurgalı Paleontoloji ve Evrim adlı kitabındaCarroll şöyle der:
Bilinen en erken amniyonluların, bu grubun üyeleri oldukları, hemen fark edilebilir. Çünkü, iskeletleri, günümüzde yaşayan gelişmemiş kertenkelelerinkine çok benzemektedir (s. 193).
İlk amniyonlulardaki en önemli damak özelliği, pterygoid’deki çapraz kenar varlığıdır.... Günümüz kertenkelelerinde, pterygoid’in çapraz kenarları, çene kapama kaslarının en büyüklerinden biri olan pterygoideus’un kökenini oluşturmaktadır.... İlkel herhangi bir amfibyumda, geniş pterigoideus kasının varlığı konusunda pek kanıt yoktur (s. 194).
İlk amniyonluların kafatasının genel yapısı, bugünkü kertenkelelerinkine benzemektedir (s. 195).
İlk amniyonluların postcranial iskeletleri, günümüz gelişmemiş sürüngen Spenodon’unkine benzemektedir (s. 195).
İlk “labyrinthodont” amfibyumlarının aksine, ilk amniyonlular, nispeten küçük kafatasları ve küçük beden ölçüleriyle, günümüzdeki gelişmemiş kertenkelelere benzemektedirler. İlk amniyonluların diş yapıları ve çene kas yapılarındaki olası düzen, neredeyse sadece böcekler ve diğer küçük eklem bacaklılarla beslenen kertenkelelerinkine benzemektedir (s. 198).
Protorothyrid’lerin iskelet anatomileri, Hylonomus ve Paleothyris, Protorothyridae familyasında yer alır Alt Pennsilvaniyen’den başlayıp, Alt Permiyen dönemine kadar nispeten değişmeden kalır. Bu zaman sürecinde, fosil kaydında, bir dizi başka amniyonlu grup ortaya çıkar. Bu grupların her biri, proterothyrid modelinden bağımsız olarak evrimleşmiş olabilir. Hiç birisi bir diğeriyle yakın bir ilişki göstermemektedir (s. 201).
Tüm Paleozoik amfibyumlarından yeterince bağımsız oldukları için, ilk amniyonluların esas ataları belirlenememiştir (s. 198).
Son durum, şüphesiz, amfibyumlar ve sürüngenler arası geçiş formlarının var olmadığını aktarmaktadır. Eğer bulunmuş olsalardı, sürüngenlerin hangi amfibyumdan geldiği ile ilgili belirsizlik de olmayacaktı.
Yukarıda da açıklandığı gibi, geçiş formlarına ne zaman ihtiyaç duyulsa bulunamamıştır. Bunun böyle olduğu, sürüngenlerin birbirinden farklı türlerinin kökenleri belgelendikçe kitabın ilerleyen bölümlerinde bol miktarda gösterilecektir. Birbirinden çok farklı bu sürüngen grupları arasında, uçan sürüngenler, deniz sürüngenleri, süzülen sürüngenler, yılanlar ve kaplumbağaları sayabiliriz.
Uçan Sürüngenler
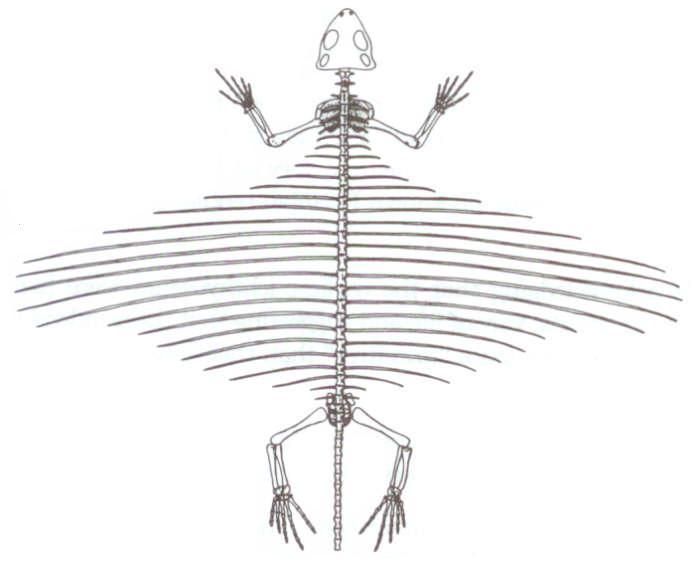
Uçan ve uçamayan sürüngenler arasında etkileyici farklar vardır. Şekil 4’te, Romer’in, kuşlar, dinozorlar ve uçan sürüngenlerin atası olduğuna inandığı tekodont sürüngenin, Saltoposuchus’un iç yapısı gösterilmiştir. Bu canlı ile, pterosaur’ların (uçan sürüngen) iki alt takımının örnekleri arasında büyük boşluk olduğu Şekil 5 ve 6’da gayet açıktır.
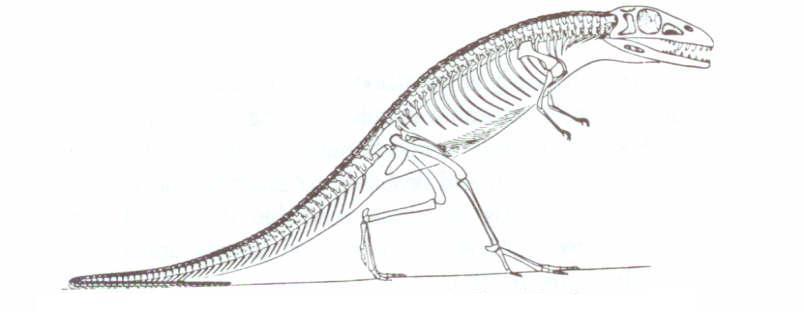 Şekil 4:Saltoposuchus (ya da Saltoposukus), Romer’in ve diğerlerinin, uçan sürüngenlerin, kuşların ve dinozorların atası olduğuna inandığı yuvaya bağlı dişli bir tekodont sürüngeni. Romer’in, Vertebrate Paleontology adlı kitabından,
Şekil 4:Saltoposuchus (ya da Saltoposukus), Romer’in ve diğerlerinin, uçan sürüngenlerin, kuşların ve dinozorların atası olduğuna inandığı yuvaya bağlı dişli bir tekodont sürüngeni. Romer’in, Vertebrate Paleontology adlı kitabından,
Chicago Üniversitesi Yayınları’nın izniyle.
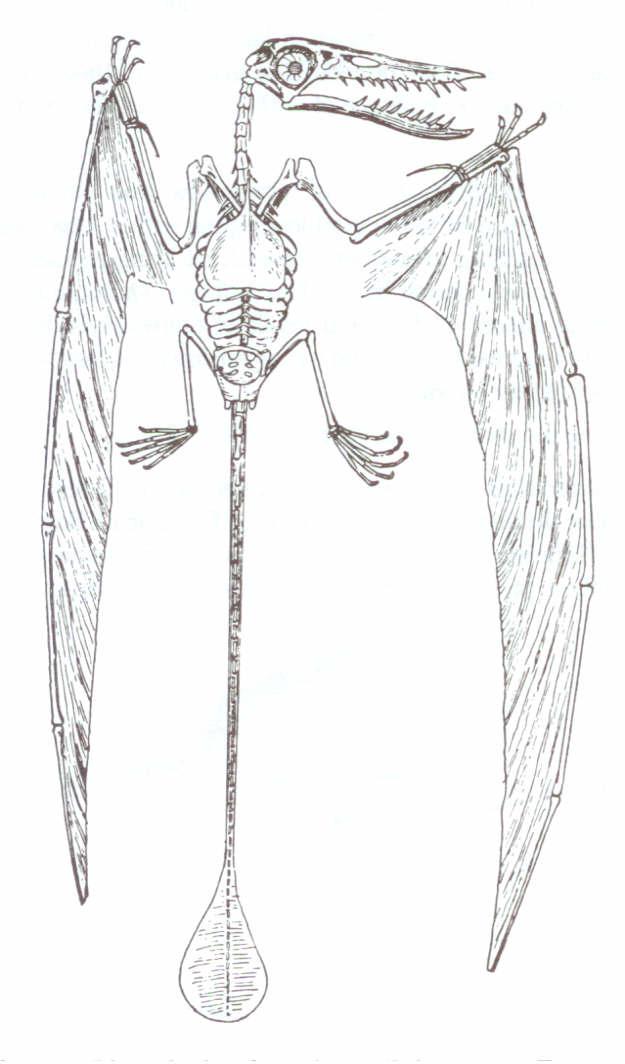
Şekil 5: Rhamphorhynchus(“Ramforhankus”), uzun kuyruklu
bir pterosaur. Williston’un The Osteology of the Reptiles
(Sürüngenlerin Kemik Yapısı) kitabından, Harvard Üniversitesi Yayınları’nın izniyle.
Uzun kuyruklu bir pterosaur olan Rhamphorhynchus’taki hemen hemen her yapı, yalnızca bu canlıya özeldi (şekil 5). Tüm pterosaurlarda olduğu gibi, sürüngende var olan üç parmağın aksine, dördüncüsünün çok daha uzun olduğu çok açıktı. Bu dördüncü parmak, kanat zarı için en büyük desteği sağlıyordu. Tabi ki bu, hassas bir yapı değildi ve eğer pterosaurlar, tekodontlardan ya da başka toprağa bağlı sürüngenlerden evrimleşmiş olsalardı, bu dördüncü parmağın kademeli olarak uzamasını gösteren ara türler bulunmuş olmalıydı. Ancak, böyle geçiş formların tek bir işaretine bile rastlanmamıştır. “En erken” pterosaurlar, Üst Trias kayaları arasında bulunmuş ve bunların, geç Rhamphorhynchus’lara çok yakın benzerlikler içerdiği açıklanmıştır.
En erken pterosaur iskeletleri, yapıları, kuşlarınkinden çok farklı olmasına rağmen, günümüz kuşlarınınki kadar etkin uçmaya uygundu. Mantığa göre, eğer, Rhamphorhynchus ve benzeri uçan canlılarda bulunan ve uçuş için gerekli olan çok özel yapıları milyonlarca yıl süren kademeli bir evrim sonucunda sıradan bir kara sürüngeninden evrimleşmişse, bu dönüşümü belgeleyen önemli sayıda geçiş formu bulunmuş olmalıdır. Böyle tek bir geçiş formu bile hiçbir zaman bulunamamıştır. Carroll şöyle der:
....bütün Trias pterosaurları uçmak için yüksek derecede özelleşmişlerdi ve pek çok özellik açısından geç rhamphorhynchus’lara benziyorlardı. Bunlar, kendilerinin esas ataları hakkında çok az, uçmanın kökeniyle ilgili erken dönemler konusunda ise hiçbir kanıt sağlamazlar.29
Pterosaurların, pterodactyl grubu da aynı derecede benzersizdi. Pteranodon (Şekil 6), büyük bir gagası ve arkaya doğru uzayan kemikli bir ibiği olan dişsiz bir sürüngendi. Bütün uçan sürüngenlerde olduğu gibi, kanat zarı, dördüncü parmak tarafından destekleniyordu. Quetzalcoatlus’un kanat açıklığı, hemen hemen 12 metreye ulaşıyordu ve Texas’ta keşfedilen pterodactyl fosillerinin 16 metreye kadar kanat açıklıklarına sahip oldukları açıklanmıştır. Bu kanat açıklığı günümüz savaş uçaklarınınkinden daha uzundur!
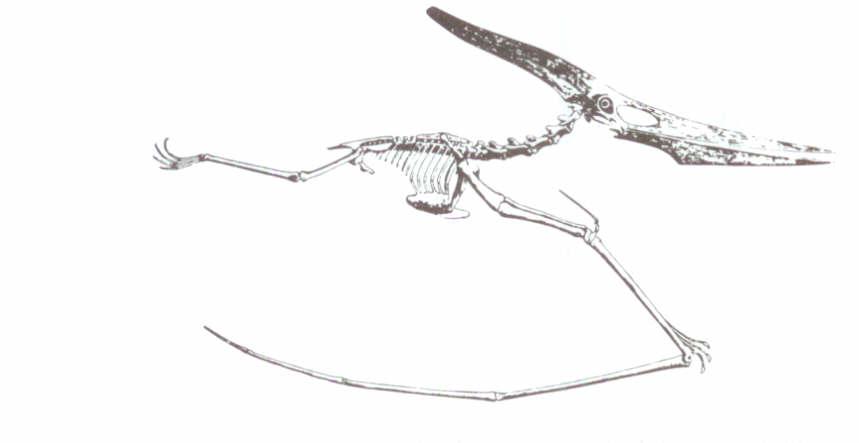
Şekil 6: Pteranodon, 15 metreden daha fazla kanat açıklığına sahip, dev bir uçan sürüngen. Romer’in, Vertebrate Paleontologyadlı kitabından, Chicago Üniversitesi Yayınları’nın izniyle.
Terodaktillerle ilgili olarak, Carroll şöyle der:
Solnhofen (Wellnhofer, 1970) fosillerinden görebildiğimiz gibi, pterodactyl benzeri canlılar, Üst Jura’da çok sayıda ve çok çeşitli olarak bulunuyorlardı. Bu kalıntılar, en küçük pterosaurların bazılarını ve en büyük uçan sürüngenlerin atalarını içermektedir. Rhamphorhynchus benzerleri ile pterodactyl benzerileri arasında bir ara form bilinmemektedir.... Uçan sürüngenlerin fosil kaydı, yaklaşık 150 milyon yıllık bir süreç ve Antarktika dışında her kıtadan alınmış yaklaşık 90 türü kapsamaktadır.30
Şu gayet açıktır ki, eğer bu çok özel uçan sürüngenler olan rhamphorhynchus benzerleri ve pterodactyl benzerleri, 150 milyon yıllık bir süreç içerisinde, bir kara sürüngeninden evrimleşmişler ve fosilleri dünyanın her yerinde bulunmuşsa, bu canlıların, bir kara sürüngeninden evrimleştiğini gösteren çok sayıda geçiş formu da keşfedilmiş olmalıdır. Durum kesinlikle tam terstir. Rhamphorhynchus ve pterodactylçeşitleri, hiçbir geçiş formu olmaksızın, tamamen gelişmiş biçimde bulunmaktadırlar.
Bir kara sürüngeninin, kademeli olarak, bir uçan sürüngene dönüşebileceği fikri, saçmalıktır. Evrimleşmeye başlamış yapılar, ara seviyelere yarar sağlamaktan çok, büyük zarar getirirdi. Örneğin, evrimciler, görülebileceği gibi, gerçekleşen mutasyonların, sadece dördüncü parmağı aşamalı olarak etkilediğini garip bir şekilde kabul etmektedirler. Tabi ki, aynı zamanda gerçekleşen, göründükleri kadar inanılmaz başka tesadüfi mutasyonlar da, kanat zarının kademeli oluşumundan, uçuş kaslarından, sinirlerden, kan damarlarından ve kanatların oluşumu için gerekli olan diğer yapılardan sorumludurlar. Uçan sürüngenin gelişimi sürecinde, sürüngen bir süre %25’lik kanatlara sahip olacaktır. Ancak, bu garip canlının yaşamını devam ettirmesine asla olanak yoktur. %25’lik kanatların ne yararı olacaktır? Açıkçası, canlı uçamayacaktır ve artık koşamayacaktır; bu işe yaramayan uzantıları ancak halen işlevsel düşünülen arka bacakları aracılığıyla sürüklemek zorunda kalacaktır. Canlı artık ne av yakalayabilecek, ne de yırtıcı hayvanlardan kaçabilecektir. Elveda dostum! Bu eşsiz uçan sürüngenlerin geçiş formları ya da atasal izlerinin en ufak kanıtının bulunamayışı, böyle bir denemeden hiç geçmediklerini göstermektedir.
Deniz Sürüngenleri
Deniz sürüngenlerinin fosil kaydında tamamıyla gelişmiş olarak aniden ortaya çıkışları, uçan sürüngenlerde karşılaşılan durum kadar etkileyicidir. Bununla beraber, evrim kuramına göre, bir kara sürüngeninin, bir deniz sürüngenine (ya da bir tatlı su sürüngenine) dönüşümü, kol ve bacakların bir küreğe dönüşümü ve ichthyosaurus’larda olduğu gibi, bir kara sürüngeninin, biçimsel ve işlevsel olarak balık benzeri bir sürüngene dönüşümü, milyonlarca yıl süren kademeli bir süreçtir. Colbert ve Morales’in bu inanılmaz süreç hakkında söyledikleri şunlardır:
Bu uzun evrim tarihi süresince, dört ayaklılar, hayatlarını tamamıyla karada geçiren canlılara, yani, sürüngenlere dönüşebilmek için, kendilerini suya bağımlı olmaktan kurtardılar. Bunlardan bazıları suya geri döndü ve sürüngenleri karada yaşayacak kadar bağımsız ve yeterli hale getiren bütün uyum türlerini değiştirmek zorunda kaldı.... Onlar, balık atalarının milyonlarca yıl önce mücadele ettiği sudaki kaldırma kuvveti, itici kuvvet ve karadan uzak üreme gibi eski problemlerini üzerlerine aldılar. Sürüngenler suya geri döndüklerinde, akciğerleri kaybolmamış ve uzun zamandan beri kayıp olan solungaçların yerine, nefes almak için kullanılmıştır. Sürüngenlerin, bacakları ve ayakları vardı. Bunlar, balıktaki yüzgeçlerle benzer işlevleri gören küreklere dönüştüler. Balık kuyruğu çoktan yok olduğu için, bazı sualtı sürüngenleri, itici kuvvet olarak, balık kuyruklarının yerine, onlara şaşırtıcı derecede benzeyen kuyruklar geliştirmişlerdir.
....Böylece bu sürüngenler arasında evrim yönünde, uzak sualtı balık atalarından, karada yaşayan ara seviye amfibyum ve sürüngen atalarına, oradan da sualtı sürüngen torunlarına doğru tersine bir dönüş olduğu açıkça görülebilmektedir .31
Bu hikaye ya da senaryoları böylesine inanılmaz yapan şey, evrimcilerin, gerekli olan şeylerin, hiçbir özel amaç gütmeyen, tamamen tesadüfi mutasyonlar ya da genetik değişimler yoluyla üretileceğine olan inançlarıdır. Sıradan kara sürüngenleri, sözde milyonlarca yıldır başarılı oldukları kara yaşamını bırakarak suya girdiklerinde, balık benzeri kuyruklara ihtiyaç duymuşlardır. Bir ihtiyaç olduğu bilinci olanaksızken, rastlantısal mutasyonlar sürüngenleri üreten karmaşık genetik yapıyı öylesine değiştirmiştir ki, önceden suda acemice debelenen sürüngenlere, güzel tasarlanmış, hayret veren bir işlevselliğe sahip, balık kuyruğuna benzeyen kuyruklar kazandırmıştır. Benzer bir şekilde, suda artık itici kuvvet sağlayamayan bacaklar ve ayakların kodlandığı karmaşık gen yapısı, bir şekilde, gelişigüzel mutasyonlar yoluyla ve mucizevi olarak, kirişlerin, kan damarlarının, sinir sistemlerinin, kasların, kemiklerin ve sudaki itici kuvvet için gerekli olan kürekleri kusursuz şekilde meydana getiren diğer yapıların kodlandığı çok karmaşık genlere dönüşmüştür. Gayet açıktır ki, ateşli yalanlamalara rağmen evrimciler, mucizelere bile inanmaktadırlar.
Bugün deniz ya da su kaplumbağaları gibi tamamıyla suda yaşayan sürüngenler geçmişte de vardı. Özellikle burada soyu tükenmiş olan plesiosaur ve ichthyosaurus gibi yaratıklara dikkat çekmek istiyoruz. Evrimcilere göre, en eski su altı sürüngeni olan ve aslında bilinen en eski sürüngen olan Mesosaurus, Alt Permiyen kayaları arasında bulunmuştur. Evrimcileri şaşırtan olay, bu canlının kol ve bacaklarının yerinde geniş perdeler bulunması ve yüzmek için uzun bir kuyruğa sahip olması gibi özel amaçlı uzuvlara sahip olmasıydı. Colbert ve Morales şöyle der:
Mesosaur’ların, erken permiyen döneminde gelişen ilkel captorhinomorph’ların neslinden gelen sürüngenlerin çok eski ve bağımsız bir evrim halkasını temsil etmeleri çok olasıdır.32
Bu balık benzeri kuyruğa ve ayak ve bacaklar yerine perdelere sahip olan özel nitelikli sualtı sürüngeni, hiçbir geçiş formu izi olmaksızın, tümüyle gelişmiş biçimde ortaya çıkmıştır. Colbert ve Morales’in yapabilecekleri en iyi açıklama, bu canlının çok çabuk evrimleştiği ve bu nedenle de geçiş formu fosili bırakabilmek için yeterli fırsatın olmadığı şeklindedir. Fakat, böylesine nadir ve doğru biçimde gerçekleşen iyi mutasyonların böylesine olağanüstü bir canlıyı üretebilmeleri için tek bir nesil dizisinde yoğunlaşması milyonlarca yıl gerektirecektir. Evrimciler, en eski sualtı sürüngenleri olan bu mesosaur’ların, geride hiçbir evrimsel torun bırakmaksızın yok olduklarına inanmaktadırlar. Carroll şöyle der:
Güney Afrika ile Güney Amerika’nın, Alt Permiyen ve Karbonifer kayaları arasında, mesosaur’lardan başka hiçbir amniyonlu bulunmamaktadır. Mesosaur’lar ile sonradan gelen başka herhangi bir sualtı sürüngen grubu arasındaki ilişki konusunda hiçbir kanıt yoktur.33
Evrimcilerin, kendi evrimsel zaman çerçevelerine göre, karşı karşıya kaldıkları daha da inanılmaz duruma lütfen dikkat edin. Güney Afrika ya da Güney Amerika’da bulunan en genç sürüngenler, sıradan kara sürüngenleri değillerdi; Alt Permiyen kayaları arasında fosilleri bulunan yüksek derecede özelleşmiş sualtı sürüngenleriydi. Alt Permiyen döneminde, yerküre kıtaları tek bir kıta halinde bulunuyor, ve bu da, sürüngenler gibi hayvanların yeryüzünde göç edebilmelerini kolaylaştırıyordu. Öyleyse, sualtı sürüngenleri, niçin sadece Güney Afrika ve Güney Amerika’nın Alt Permiyen kayaları arasında bulundu; evrimsel kökenleri, sıradan bir kara sürüngeninden gelen bu canlılar için milyonlarca yıl gerekmiyor muydu? Erken sürüngenler olarak isimlendirilen bir kaç fosil, Kuzey Amerika ve Avrupa’nın Alt ve Orta Pennsilvaniyen kayaları arasında bulundu. Sözde bu canlılar, Permiyen’den milyonlarca yıl önce var olmuşlar ve bu nedenle de, Permiyen zamanına kadar Kuzey Amerika’dan Güney Amerika’ya ve Avrupa’dan Güney Afrika’ya göç edebilmek için çok fazla zamanları olmuştur. Öyleyse, erken bir kara sürüngeninden türediği düşünülen sualtı sürüngenleri, niçin sadece Güney Afrika ve Güney Amerika’nın Permiyen kayaları arasında bulundu? Niçin Güney Amerika ve Güney Afrika’nın Pennsilvaniyen ve Permiyen kayaları arasında, bu ileri derecede özelleşmiş sualtı sürüngenleri dışında başka herhangi bir çeşit amniyonlu bulunmuyor? Evrimsel senaryoların aslında hiçbir anlamı yoktur.
Şimdi, ileri derecede özelleşmiş olan plesiosaur ve ichthyosaurus’lar gibi deniz sürüngenlerini ele alalım. Jura ve Kretase kayaları arasında yer alan plesiosaur fosilleri tüm dünyada yaygındılar. Jura kayaları arasında bulunan plesiosaur fosilleri üç ile altı metre uzunluğunda, Kretase kayaları arasında bulunanlarınki ise on iki metre ve daha fazla uzunluktaydılar. Birçoğunda bu uzunluğun büyük bir kısmı uzun boyundan ibaretti. Çok geniş perdeleri vardı ve güçlü kasların konumu, perdelerini hem geri, hem de ileri doğru büyük bir güçle çekmelerini sağlıyordu. Hiç kimse, hiçbir yerde, ne ayak ve bacakların perdelere kademeli dönüşümünü gösteren geçiş formlarının fosillerini, ne de plesiosaur’ların başka birçok eşsiz özelliğinin kökenini bulabildi.
Nothosaur’lar uzun, yılansı boyunlu sualtı sürüngenlerdi. Bunlar, kısa perdeliydiler. Kuyruklarının, bu sualtı sürüngenleri için ana itici kuvvet sağladığına inanılmaktadır. Nothosaurların, plesiosaur’ların mümkün atası olabileceği ileri sürüldü. Carroll, nothosaur’ların pek çok iskelet özelliğinin, onların, plesiosaur’ların olası ataları olduğunun bir göstergesi olduğuna inanıyor. Fakat, daha sonra şöyle der:
Ancak, böylesine bir ilişki, damak ve omuz kemerlerinin yapıları ile çelişmektedir. Plesiosaur’ların damakları, nothosaur’larınkinden daha az özelleşmiştir... Bu yapı, plesiosaur’ların, herhangi bir iyi tanınmış nothosaur’dan çok, daha ilkel bir diapsidden evrimleşmiş olabileceğini düşündürmüştür.... Plesiosaur’ların nothosaur’dan geldiği kabulü, bir diğer problemi ise, omuz kemeri şekillerindeki büyük farklılıktır.34
Eğer fosilbilimciler, plesiosaur’lara öncülük eden geçiş formları bulabilmiş olsalardı, atalar ve torunlar arasındaki evrimsel yolları belirlemede hiçbir zorluk olmayacaktı. Evrimciler, bunlara sahip olmadan, yalnızca, kuramsal evrim yolları ileri sürebilmektedirler.
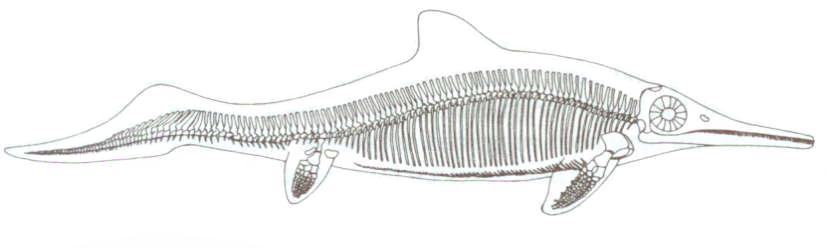
Şekil 7: Bir ichthyosaurus gösterimi.
Ichthyosaurus’lar, evrim kuramına karşı, gayet açık ve ikna edici bir çelişkiyi göstermektedir. Ichthyosaurus’lar, diğer tüm sürüngenlerden, çarpıcı biçimde farklı olan deniz sürüngenleriydi. “Balık sürüngeni” anlamına gelen ichthyosaurus’lar, balığa çok benzeyen canlılardı ve üç metre ya da daha fazla bir uzunluğa sahiplerdi. Fosilleri, dünyanın pek çok yerinde, Jura kayaları arasında bulundu ve bu fosillerde, fosilleşmiş kemiklerin yanı sıra, korunmuş bedenlerinin ana hatları da bulunmaktadır. Öyleyse, elimizdeki şey, yaratılış ve evrim kuramını birbirlerine göre değerlendirmenin çok uygun bir örneğidir. Bu inanılmaz derecede eşsiz canlı, evrimsel varsayıma göre milyonlarca yıllık bir süreçte, tesadüflere dayalı mutasyonların çıkardığı kademeli değişimler aracılığıyla çok farklı türdeki bir sürüngenden ortaya çıkmıştır. Bu kuramsal dönüşüm süresince ortaya çıkan geçiş formu dizileri, bu dönüşümün gerçekten olduğu konusunda açık ve belirli kanıtlar ortaya koyacaklardır. Farklı zamanlara ait bir kaç geçiş form, ichthyosaurus’ların iddia edilen evrimsel köken gerçeğinin açığa çıkarılabilmesi için yeterli olacaktır. Dünyanın her yerinde ichthyosaurus fosilleri bulunmuştur. Bu nedenle, geçiş formların yokluğuna açıklama getirebilmek için ortaya atılan fosillerin çok nadir oluşu, yerel ya da bölgesel şartlar ve olaylar gibi iddiaların, kesinlikle hiçbir geçerliliği yoktur. Elbette ki, milyonlarca yıla yayılmış tüm geçiş formların, fosilleşmeden ya da fosilleştikten sonradan yok olmaları ve her nasılsa sadece uç formların fosilleşerek kalması inanılmazdır. Öte yandan, eğer, ichthyosaurus’lar, fosil kaydında, hiçbir geçiş form izi olmadan tümüyle gelişmiş biçimde aniden ortaya çıkmışlarsa, bu, yaratılış için olumlu ve güçlü bir kanıt sağlayacaktır. Bu durumda gerçek nedir?
Colbert ve Morales şöyle derler:
Pek çok yönden en ileri derecede özelleşmiş deniz sürüngenleri olan ichthyosaurus’lar, erken trias döneminde ortaya çıkmıştır. Onların, sürüngenlerin jeolojik tarihçelerine girişleri, çok ani ve çarpıcıdır; Trias öncesi tortularda, ichthyosaurus’ların olası ataları konusunda hiçbir ipucu yoktur. Onların kökenleri hakkında yapabileceğimiz tahminler, ileri derecede özelleşmiş sürüngenlerin anatomik yapılarının yorumundan başka bir şey değildir. Bu köken olasılıkla erken diapsid atalardan gelmektedir. Ichthyosaurus’un akrabaları konusundaki esas sorun, bu sürüngenleri başka sürüngen takımlarına bağlayan hiçbir kesin kanıtın bulunamayışıdır.35
Romer, ichthyosaurus’ların, ilk ortaya çıktıklarında, ileri derecede özelleşmiş formlar olduklarını kabul ettikten sonra şöyle der:
Daha erken hiçbir form bilinmemektedir. Ichthyosaurusun yapısal özelliklerinin gelişimi için, görünüşe göre, uzun bir zaman ve bundan dolayı da çok erken bir köken gerekmektedir; fakat, onun kendisinden önce hiçbir Permiyen sürüngeni bilinmemektedir.36
Carroll, en erken ichthyosaurus’ların, ileri derecede suya dayalı bir yaşam biçimine uyarlanmış olduklarını kabul ederken ve bu bireylerin daha sonraki kopyalarından daha “ilkel” özelliklere sahip olduklarını ileri sürerken şöyle der:
Ancak, en ilkel özellikler bile, onlarla, belirli bir karasal ya da deniz sürüngen grubu arasında bağlantı kuramamaktadır.... 37
Böylece, sonuçlar birbiriyle uyuşmaktadır. Şekil ve hareket açısından tümüyle balığa benzeyen, denizde yaşarken yavru yapan, yeryüzünde herhangi bir zamanda var olmuş sürüngenlerin en ileri derecede özelleşmiş biçimi olan ichthyosaurus’lar, fosil kaydında, hiçbir geçiş form ya da ata izi olmaksızın, aniden, tümüyle gelişmiş biçimde ortaya çıkmıştır. Üstelik, Colbert ve Morales’in de kabul ettikleri gibi, ichthyosaurus’ları, başka herhangi bir sürüngen grubuna bağlayan hiçbir gerçek kanıt yoktur. Ichthyosaurus’lar ile diğer tüm sürüngen takımları arasında şüphesiz geniş ve doldurulamayan bir boşluk vardır. Burada, evrimcilerin, ichthyosaurus’lara en yakın oldukları iddia edilen sürüngen takımlarının çeşitli benzerlikleri konusundaki bitmez tükenmez tartışmalarını görmüyoruz; fakat burada, geçiş formları eğer gerçekten var olmuş olsalardı, varsayılan karasal ya da deniz sürüngenin kademeli olarak balık benzeri ichthyosaurus’a dönüşmelerini anlamak kolay olurdu. Doğa tarihi müzeleri, herkesin görebilmesi için, bu tür belirli geçiş formlarını sergilemeleri gerekirdi. Bu, evrim için, şüphesiz bir kanıt olurdu. Eğer kanıtlamak mümkünse, sahip olduklarımız, özel yaratılışın şüphesiz kanıtıdır. Kayalar, yaratılış için daha hangi büyük kanıtları sağlayabilirler ki?
Şekil 8: Coelurosaurus, Madagaskar’daki Permiyen kayaları arasında bulunmuş, süzülen bir sürüngen. Carroll’un, Vertebrate Paleontology and Evolution adlı kitabından. Telif hakkı, 1994, W. H. Freeman and Company’e aittir. İzinle kullanılmıştır.
Süzülen Sürüngenler
Süzülen sürüngenler, önemli mesafelere süzülebilmelerini mümkün kılan, zarla desteklenmiş, büyük ölçüde uzun kaburga kemikleri olan, ileri derecede özel sürüngenlerdi. Draco cinsi günümüz sürüngenleri, ağaçtan ağaca, neredeyse 30 metre uzaklığa süzülebilen, sürüngenlerin yaşayan temsilcileridirler. Coelurosauravus, fosilleri, üst permiyen kayaları arasında bulunan, çok sayıda uzun kaburga kemikleri olan bir süzülen sürüngendir. Fosilleri, üst Jura kayaları arasında bulunmakta olan Kuehneosaurus, bu ileri derecede özel süzülen sürüngenlerin başka bir çeşididir. Süzülen sürüngenlerin herhangi birisinin, diğerleriyle akraba olduğunu hiçbir paleontolog söylememektedir.. Carroll, kaburgaların, süzülme zarını desteklemek için, yatay yönde aşırı uzun olması konusunda şöyle der, “bu özelleşme, bu grupların her birinde ayrı ayrı evrimleşmiştir.”38 Ne Carroll, ne de Colbert ve Morales, sıradan bir kertenkelenin ya da sürüngenin süzülen bir sürüngene dönüşümünü gösteren geçiş formlarının yokluğunu ima bile etmemişlerdir. Fosil kaydında, her süzülen kertenkelenin ilk örnekleri, hiçbir türün geçiş formu olmaksızın, bütün olarak görünmektedir. Bu kanıt, evrim kuramının dayandırıldığı tahminlerle doğrudan çelişkili olmakla birlikte, yaratılış kuramının dayandırıldığı tahminlerle tam bir uyum içindedir.
Yılanlar
Yılanlar ve kertenkeleler, Squamata takımında yer alırken, kertenkeleler, Lacertilia ya da Sauria alttakımında, yılanlar da Ophidia ya da Serpentes alttakımında yer almaktadırlar. Evrimciler, yılanların, kertenkelelerden evrimleştiğini tahmin etmektedirler. Yılanlar, kuramsal olarak evrimsel ataları olan kertenkelelerden pek çok yönden farklı, ileri derecede özel nitelikli sürüngenlerdir. Yılan gövdeleri bazı yılanlarda, kuyruk önü omurları 120’den 454’e kadar değişen bir kaç yüz omura kadar uzamaktadır. Bazıları küçük bir pelvis kemerine ve arka üyelere sahip olmalarına rağmen, yılanlarda, ön üyeler ve göğüs kemeri yoktur. Bunlar, evrimcilerin inandığı gibi körelmiş yapılar değillerdir fakat, önemli işlevlere sahiptirler. Pelvis kemikleri, belirli kaslar için dayanak noktası olarak, arka üyeler ise, çiftleşme sürecinde, ve belki de hareket sırasında yüzeyi kavramak amacıyla kullanılmaktadır. Alt ve üst çene yapıları ve damak, büyük bir avı bir bütün olarak yutabilmek için ağzın geniş açılmasını sağlayacak şekilde kertenkeleden farklıdır. Avın yutulması sırasında, beyin korunmalıdır. Alın ve kafatası çeperi kemikleri, beynin tamamıyla korunacağı bir şekilde daha geniştir. Ayrıca, omurlar da yılan vücudunun kıvrak hareketine imkan vermek için değişiktir.
Evrim kuramına dayanarak, bir kertenkelenin, bir yılana dönüşümü sırasında gerçekleşen ve bu dönüşümü belgeleyen kafatası değişimleri, ön ve arka üye kayıpları, omurların kademeli olarak çoğalması ve buna benzer başka önemli değişimler ümit edilecektir. Keza, şu soru cevaplandırılmalıdır: olağanüstü başarılı bir kertenkele herhangi bir seçilim yoluyla niçin ön ve arka üyelerin kaybıyla sonuçlanacak ve kendisinin çok yetenekli koşma kabiliyetini, yetersizliği kesin, kertenkele ve yılan arası bir düzeye dönüştürecek olan tedrici değişimlere maruz kalsın? Kertenkelenin yılana evrimi konusundaki fosil kanıtları hakkında ne söylenebilir? Yeterli kafatası kalıntısı bulunan en eski yılan fosili, Dinilysia, Güney Amerika’nın Üst Kretase kayaları arasında bulundu. Carroll, Dinilysia hakkında şöyle der: “bu özellikler, oldukça ilerlemiş bir evrim seviyesini gösteriyor” yılan kafatasının farklılaşmış özellikleri.39 Colbert ve Morales şöyle der:
Ne yazık ki, yılanların fosil tarihi çok eksik kalmıştır; bu yüzden, onların evriminin birçoğunu modern biçimlerinin karşılaştırmalı anatomisinden çıkarmak gereklidir.40
Romer, yılanların fosil yönünden çok fakir olduklarını ifade ettikten sonra şöyle der:
Yılanlar, kesinlikle kertenkelelerden türemişlerdir ve başlangıç noktasıyla ilgili hiçbir kesinlik yokken, varanoid'lerle akrabalıkları oldukça olasıdır....41
Eğer yılanlar, bazılarının inandığı gibi, gerçekten varanoid kertenkelelerden evrimleşmişlerse, kertenkelenin yılana kademeli dönüşümünü belgeleyen yeterli sayıda fosil kaydı da olmalıdır. İddia edilen kertenkele atasının ve yılanların binlerce fosili bulunmaktadır. Eğer, sadece bir kaç tane kertenkeleyılan arası ara seviye form bulunabilmiş olsaydı, ata kertenkeleye ya da ata olduğu kanıtlanan sürüngene doğru iz sürerek, yılanların kökenini araştırıp bulabilmek mümkün olacaktı. Colbert ve Morales’in de itiraf ettikleri gibi, geçiş formları bulunamadığı için, varsayılan evrimsel “tarih”in çoğu, canlı formların karşılaştırmalı anatomilerine bakılarak tahmin edilmek zorunda kalınmıştır.
Kaplumbağalar
Kaplumbağalar, tarih boyunca var olan, en ileri derecedeki özel sürüngenlerdendir. Kaplumbağaların eşsiz, kemikli, bağa ya da karapaks denen koruyucu bir kabuğa sahip olmalarının yanı sıra, dışta ya da sırtta olan kaburgaların yerleri ile içeride ya da karında yer alan göğüs ve pelvis kemerinin yerleri ters durmaktadır. Kaplumbağalar inanılmaz derecede eşsiz bir yapıya sahiptirler, bundan dolayı kaplumbağalar, bir çeşit sıradan sürüngenden evrimleşmiş olsalardı, ata sürüngenden kaplumbağaya giden evrimsel yolları gösterecek geçiş formlarını anlamak çok kolay olacaktı. Gereken değişimler, gizli değil, anatomi ve paleontoloji eğitimi gerektirmeyecek kadar açık olacaklardır. Evrimciler arasında, hangi sürüngenin gerçek ata olduğu ve hangi fosilleşmiş canlıların ara seviyeleri oluşturduğu konusunda hiçbir tartışma olmamalıdır. Diğer yandan, yaratılışçıların tahminlerine göre, kaplumbağaların, fosil kaydında ilk ortaya çıkışlarında, tümüyle gelişmiş bağaya ve diğer eşsiz özelliklere bakılarak kaplumbağa oldukları hemen fark edilebilir olmalıdır. Kaplumbağa fosil kaydını hangi köken modeli desteklemektedir? Yaratılış mı, yoksa evrim mi?
Romer şöyle der:
Takımın fosil üyeleri çalışmalarıyla, kaplumbağaların esas kökeni konusuna pek az ışık tutmuştur. Proganochelys gibi bazı erken Trias formları, sonraki kaplumbağalarınkinden biraz daha ilkel bir yapı göstermiştir. Fakat o zamanda bile zırh, hemen hemen mükemmel bir biçimde gelişmişti; biz, bir geçiş formundan değil, kesinlikle gerçek bir kaplumbağadan bahsediyoruz.42
Colbert ve Morales şöyle yazarlar:
İlk gerçek kaplumbağalar, tipik günümüz kaplumbağalarının yayılma yolu ile geliştikleri bir zaman olan trias döneminin son bölümlerinde ortaya çıkmışlardır. Karakteristik bir Trias cinsi olan Proganochelys’de, kafatası kemikleri sayıca azalmış, çene kenarından itibaren dişler yok olmuş ve gövde, ağır bir kabukla korunmuştu. Bunlar, temel kaplumbağa adaptasyonlarıdır ve Trias zamanından beri kaplumbağalar arasında gerçekleşen evrim, genel anlamda, Proganochelys’de ortaya çıkan karakteristiklerin rafine edilmesine yönelik olmuştur.43
Carroll şöyle der:
En erken kaplumbağalar, Almanya’daki Üst Trias tortuları arasında bulunmuştur. Bu kaplumbağalar, günümüz cinsleriyle yakın olan kabuk şekillerinden hemen tanınabilmiştir. Kaplumbağa kabukları, kolayca fosilleşebilen ve en küçük parçası bile kolayca tanınabilen kabuklar olmalarına rağmen, erken ya da daha ilkel kaplumbağalar hakkında hiçbir iz tanımlanamamıştır.
Kabukların evrimiyle ilgili olarak, en erken kaplumbağaların postcranial iskeletleri bile ilkel amniyonlularından öylesine başkalaşmışlardır ki, bu canlıların esas akrabaları konusunda pek az ipucu vermektedir. Ayrıca, geç trias dönemi kaplumbağalarının kafatasları da ileri derecede özelleşmiştir. Erken sinapsidler ve diapsidlerle yakın akrabalığı imkansız hale getirecek şakak açıklığının önceki varlığı konusunda hiçbir kanıt yoktur. Sonradan oluşmuş geniş açıklıkların varlığı ve damaktaki ectoterygoid kemiklerin yokluğu, Captorhinidae Familyası ile paylaşılan özelliklerdir. Fakat, diğer yandan, kafatası öylesine değişmiştir ki, bizler, kaplumbağaların, erken anapsidlerin soyu olan bu familyanın yerine başka bir anapsid familyasından evrimleştiğini güvenle ortaya koyamayız.44
Gayet açıktır ki, çok özel sürüngen olan kaplumbağaların geçiş formları, eğer aslında var olmuşlarsa, bol miktarda ve kolayca fark edilebilen olmalıdır. Fakat, tek bir geçiş formu bile keşfedilmemiştir.
Son yıllarda, Michael S. Y. Lee, “Kaplumbağa Vücut Şeklinin Kökeni: Ünlü bir Morfolojik Uçurumun Kapanması” (The Origin of the Turtle Body Plan: Bridging a Famous Morphological Gap) adlı bir makale yayınladı45. İnsanlar, bu başlığa bakınca, kaplumbağaların, kafatası ya da kaburga kemeri ve kafesinin tersine dönüşü gibi eşsiz karakterlerinin başlangıç safhasındaki henüz tanımlanmamış geçiş formlarından bazılarının fosilleşmiş kalıntılarının keşfi konusunda bir hamle yapıldığını düşünebilirler. Lee’nin yazısının içine aldığı şey, yanak kertenkelesinin(pareiasaur) yeni bir incelemesinden başka bir şey değildir. Lee’ye göre, kaplumbağa ve pareisaur karakteristikleri konusundaki çalışmalar, pareisaur’ların, ilkel sürüngenler arasındaki en yakın kaplumbağalarla akraba olduğuna karar verilmesine yetecek kadar çok benzerlik içermektedirler. Peki, pareisaur’lar neydi? Onlar, üç metre uzunluğunda, büyük, hantal sürüngenlerdi. Ön ve arka üyeleri, kısa ve kalındı ve ağır bir gövdeyi taşıyorlardı. Carroll onları “fil gibi hayvanlar” olarak isimlendirir. Tabi ki, onların fosillerine bir göz atmak, onları, kaplumbağalara en ufak bir şekilde bile benzemeyen kaplumbağaların en az olası atası olarak nitelendirmeye yetecekti. Lee’nin, pareisaurları, kaplumbağalarla birleştirme girişimleri, kaplumbağalarla başka sürüngenler arasındaki olası ata boşluğunun kapanmadığını daha da çok kanıtlamıştır.
Böylece, görüyoruz ki, sürüngenler grubuna giren uçan sürüngenler, deniz sürüngenleri, süzülen sürüngenler, yılanlar ve kaplumbağalar gibi eşsiz ve ileri derecede özel olan canlıların dikkatli bir incelemesi, bu özel sürüngenler ile diğer tüm sürüngen takımları arası geçiş formlarının sistematik boşluğunu açığa çıkarmaktadır. Geçiş formlarına en çok ihtiyaç duyulan ve geçiş formlarının fark edilebilmesi en kolay olan bu aralarda, onlar hiç yoktur. Bu gerçekler, özel yaratılış için, güçlü ve olumlu bir kanıt sağlamaktadır.
Dinozorlar – Yaratılış İçin Güçlü Bir Kanıt
Dinozorlar, şimdiye kadar yaşamış canlılar içinde, insanları ve özellikle de çocukları en çok büyüleyen canlılardır. Bunun nedeni belki de onların, birçok örneğinde görülen büyüklükleri ve sahip oldukları pek çok alışılmadık anatomik özellikleridir (dinozorların büyüklüğü, bir horozun boyutundan seksen tona kadar bir ağırlığı ve beş katlı bir binaya kadar uzunluğu olan Brachiosaur’unboyutuna kadar değişkenlik gösteriyorlardı). Dinozorların fosil kaydı, bugün soyları tükenmiş olan canlılar konusunda mümkün olduğu kadar yaratılışı doğrulamaktadır.
İlk dinozor fosili, 180 yıl önce İngiltere’de bulundu. 1822 yılında, bir fizikçi ve aynı zamanda da amatör bir paleontolog olan Dr. Gideon Mantell’in eşi tarafından büyük bir diş keşfedildi. Dr. Mantell, daha başka fosiller aradı ve birkaç diş ve kemik daha buldu. Bulduğu fosilleri ünlü bir Fransız bilim adamı olan Baron Cuvier’e gönderdi. Şimdiye kadar, bunlarla karşılaştırılabilecek hiçbir fosil görmemiş olan büyük Fransız bilim adamı bu kez yanıldı. Cuvier, dişlerin, eski bir gergedana ve kemiklerin de, soyu tükenmiş bir suaygırına ait olduğunu belirledi.
Daha sonra, Dr. Mantell’in bir arkadaşı, ona, dişlerin yapısının, Meksika ve Güney Amerika’da bulunan Hint (İguana) kertenkelesininkilere benzediğini haber verdi. O zaman Dr. Mantell, çok şaşırtıcı yeni bir canlı türünün kalıntılarını bulduğuna karar verdi ve bu yaratığa, Iguanodon (iguana dişli) adını verdi. Kısa bir süre sonra, kertenkele benzeri büyük bir etçilin fosilleşmiş diş ve kemikleri bulundu ve Megalosaurus olarak isimlendirildi. Iguanodon ve Megalosaurus’un, o güne kadar keşfedilmemiş, soyu tükenmiş bir canlı türünün üyeleri oldukları fark edildi. Büyük İngiliz anatomi ve fosil bilimcisi Sir Richard Owen, onlara, “korkunç kertenkele” anlamına gelen Dinosaur ismini verdi.
Iguanodon kalıntılarıyla birlikte, keskin, gaga benzeri bir kemik bulundu ve bunun bir gaga olduğu düşünüldü. Ancak, 1877’de, Beçlika’da kömür madeninde “gaga”nın gerçek doğasını ortaya çıkartan, harika bir keşif yapıldı. Madenciler, kömür madeninin derinliğinde, yaklaşık 25 tane fosilleşmiş Iguanodon iskeleti buldu. Kömür madeninin derinlerinde böyle şaşırtıcı bir Iguanodon fosil mezarlığı bulunması, evrimci jeologlar için zor açıklanabilen bir durum ortaya koymuştur. Bazıları, bir zamanlar, yeryüzünde, Iguanodon'ların yuvarlanıp öldükleri derin bir çatlak olduğunu savundu. Ancak, diğer canlıların bu tuzaktan kurtulmayı nasıl başardıkları da çok tuhaf görünüyor. Sadece bir kaç kişi, bu canlıların ve gömülüp kömürleşmiş bitki materyallerinin büyük bir su afetiyle oraya sürüklendiğini ve örtüldüklerini ileri sürmüştür.
Fosil çalışmaları, “gaga”nın, gerçekte bir başparmağa benzediğini gösterdi. Iguanodon'ların, “başparmaklarını” nasıl kullandıklarını bilmek imkansızdır. Belki de başparmak, bitkileri soymak için kulanılıyordu. Bazıları bunun, savunma için kullanıldığını önerdi.
Sürüngenimsi ve kuşsu dinozorlar (Saurischia ve Ornithischia Takımları), timsahlar (Crocodilia Takımı), uçan sürüngenler (Pterosauria Takımı) ve başka çeşit sürüngenler (Tekodontia Takımı), tümünün sahip oldukları pek çok ayırt edici özellik yokken ve böylece birbirine bağlayan pek bir şey olmamalarına rağmen bu takımlar yine de Archosauria (egemen sürüngenler) alt sınıfı altında birlikte gruplandırıldılar. Saurischia dinozorlarını, ornithischia dinozorlarından keskin bir biçimde ayıran ayırt edici özellikler vardır. Ornithischia dinozorları “kuş benzeri” kalçalara sahipken, saurischia dinozorları, “sürüngen benzeri” pelvis ya da kalçaya sahiptirler. (Bu iki pelvis yapısının açıklaması için: Dinozorlar: Tanrı’nın Olağanüstü Tasarımı, Yeni Yaşam Yayınları, İstanbul, 2005, s. 65). Ayrıca, kuşsu dinozorlar, sürüngenimsilerde bulunmayan bir predentary kemiğe sahiptiler. Birçok kişi tarafından, tekodontların ve özellikle Thecodontia alttakımı Pseudosuchia’ların, timsahların, dinozorların, uçan sürüngenlerin ve kuşların ataları olan sürüngenler oldukları düşünülmektedir.
Bu varsayımsal atanın uydurulmuş olduğu, evrim literatüründe birdenbire ortaya çıkmaktadır. Romer, bir pseuodosuchia tekodont’u olan Saltoposuchus (bkz: şekil 4) hakkında konuşurken şöyle der:
Bunun, pterosaur’lar, kuşlar ve dinozorların ortaya çıktığı türe ait bir form olduğu gayet açıktır. Ne sözü edilen ilk iki gruba, ne de iki dinozor takımından birine, Ornithischia’ya, doğru olumlu bir işaret gösteren hiçbir bilinen tekodont yoktur.46
Eğer bu canlılar, uçan sürüngenler, kuşlar ve ornithischia’lara doğru hiçbir “olumlu işaret” göstermiyorlarsa, Saltoposuchus gibi bir canlının, nasıl oluyor da uçan sürüngenlerin, kuşların ve kuşsu dinozorların atası olduğu gayet açık oluyor? Görünen o ki, Romer, daha iyi bir adaydan yoksun olduğu için, kuşların, uçan sürüngenlerin, kuş kalçalı dinozorların ve timsahların ataları olarak tekodont sürüngenleri seçmiştir. Çünkü, fosil kaydı, gerekli geçiş formlarını ve esas ataları göstermemektedir.
Romer, archosaur’lar (tekodontlar, timsahlar, uçan sürüngenler ve dinozorlar) konusundaki önceki tartışmasında şöyle der:
Farklı archosaur’ların uç formları arasındaki yapısal özelliklerdeki pek çok benzerlik, ortak bir atadan gelir biçimde, bundan sonraki döllere aktarılmadı. Fakat, bu özellikler, farklı grupların üyeleri tarafından bağımsız olarak kazanıldılar. Ancak bu, akrabalık göstergeleri olarak karakterlere bir engel teşkil etmez. Fosil form çalışmaları gittikçe artan bir şekilde göstermektedir ki, evrimde, çok büyük miktarda bir paralellik vardır; fakat, yine bu çalışmalar, yakın paralelliklerin, yalnızca, yakın akraba formlarında gerçekleşmiş gibi görünmektedir.47
Romer’in burada kabul ettiği şey, bu canlı gruplarının, pek çok benzerlik içermeleri nedeniyle birbirlerine bağlandıkları, fakat her durumda, yapısal özelliklerin pek çoğunun ve varsayılan evrimsel uç formları tarafından ortaklaşa paylaşılmayan özelliklerin zaten varsayılan ortak ata tarafından da sahip olunmadığı ve böylece sonraki döllere de aktarılmadığıdır. Evrimcilerin bu gibi durumlarda kabul ettikleri şey, evrimsel ata popülasyonunun, ayrı ayrı evrimleşen soylara bölünmesiyle, her evrimleşen soyun, aynı yapı ya da yapıları diğerlerinden bağımsız anlamda kazandıklarıdır (“paralel evrim”). Bunun anlamı şudur: Bir yapısal özelliğe ortak anlamda sahip olan iki veya daha fazla canlının, eğer, varsayılan atanın, bu yapısal özelliği taşımadığı biliniyorsa, sahip oldukları bu özellik, onların ortak bir atadan geldiklerini göstermemektedir.
Romer, timsahlar, uçan sürüngenler, tekodont’lar ve dinozorların, sahip oldukları benzerlikler nedeniyle Archosauria adlı tek bir grupta toplandıklarını söylemekte ve bu benzerliklerin birçoğunun ortak bir atadan aktarılmadığını söylemeye de devam etmektedir. Bununla beraber, bu benzerlikler, ataların tahmin edilmesinde bir temel görev görmektedir. Romer, yakın paralelliğin, yalnızca yakın akraba formlarda meydana geldiği tahminine dayanarak, evrim masalını haklı çıkarmaya çalışmaktadır. Fakat, eğer benzerlikler, bağımsız olarak kazanılmış olan akrabalıkların ortaya çıkarılmasında kullanılıyorsa, o zaman, sorgulanan canlıların gerçekten akraba olduklarını nereden bileceğiz? Bu gerçekten de kör bir inanç gerektirir.
“Erken Dinozor Evriminin Sırları” başlıklı bir makalede Cox şöyle der, “Dinozorların soylarının tükenmesinin sırrı konusunu tartışan çok şey yazılmaktadır, bu sırrın bir o kadarı da, onların kökeni ya da kökenlerini kuşatmış olan belirsizliktir.48
Önceden de belirtildiği gibi, dinozorların, pseudosuchia’lardan evrimleştiği tahmin edilmektedir. Bu fikirdeki sorun, ata kabul edilen pseudosuchia’ların, dinozorlar evrimleştikten sonra, halen var olmalarıdır. Cox şöyle der:
Geç Trias’ın her döneminde, pseudosuchia’lar, torunları oldukları tahmin edilen büyük çeşitlilikteki dinozorla beraber ve büyük bir çeşitlilikle, bir arada var olmuşlardır.
Cox, kuşsu ve sürüngenimsi dinozorların hakkında konuşurken şöyle der:
Eğer Trias pseudosuchia’ları ile bu iki çeşit dinozor arasında bir bağlantı kurulmaya çalışılırsa, aralarındaki olası evrimsel bağların inatla ortaya çıkmamasına rağmen, bu iki grup arasında zaman olarak şaşırtıcı bir çakışma var olduğu görünmektedir.
Önceden de belirtildiği gibi, Romer, kuşsu dinozorları, tahmin edilen tekodont atalarıyla bir araya getirmenin hiçbir yolu olmadığını kabul etmektedir. Ancak, Cox’un da anlattığı gibi, her ne kadar, pseudosuchia tekodont atalar ve onların dinozor dölleri Geç Trias’ta bir arada var olmuşlar ve onların arasında hiçbir evrimsel ara tür bulunamamışsa da, Romer, sürüngenimsi dinozorların tekodontlara benzerliklerinin, onların tekodont sürüngenlerden evrimleştiğini düşünmeye yetecek kadar büyük olduğunu kabul etmektedir.
Kökenler ile ilgili yaratılış ve evrim modellerinin güvenilirliği konusunda düşünürken, dinozorların esas kökeni konusu ile ilgili olarak, en doğru yaklaşım, bir tekodont sürüngeni ile genel bir dinozor formu arasında bir bağ hayal etmek değil, benzersiz yapılara sahip dinozorların evrimsel kökenlerini belgeleyecek olası geçiş formlarını aramaktır. Eğer evrim gerçekse, bu eşsiz yapıların kademeli olarak var oluşlarını gösteren bir dizi geçiş formlarını kolaylıkla bulabilmeliyiz. Eğer yaratılış gerçek ise, bu eşsiz yapılara sahip dinozorlar, başlangıçta, tümüyle gelişmiş biçimde, birdenbire ortaya çıkacaklardır. Bu durumda yaratılış kuramı kesin olarak kazanmış oluyor. Gerekli olan geçiş formları tek bir durumda bile bulunmamıştır.
Boynuzlu dinozorların (Ceratopsia altsınıfı) en önemli özelliği, tabi ki, bir veya birden daha çok boynuzları olmasıdır. Bu kuşsu dinozorların kemikli boynuzları, görünüş olarak, günümüz bizonlarınınkine benzemektedir. Triceratops, yaklaşık sekiz ile on tonluk bir ağırlığa ve beş ile altı metre arasında bir uzunluğa sahipti. Her iki gözünün üzerinde birer tane ve burun bölgesinde de bir tane merkez boynuz olmak üzere toplam üç büyük boynuza sahipti. Triceratops, kafatasının, parietal ve squamosal kemiklerinin uzamasıyla oluşmuş, onon beş santim kalınlığında, kemikli bir boyun yelesine sahiptir. Bu kalkan, boyun bölgesi için önemli bir koruma sağlıyordu. Centrosaurus da, kemikli bir kalkanla donatılmıştı ve burun bölgesinde tek bir boynuza sahipti.
Protoceratops, Moğolistan’ın Üst Kretase kayaları arasında bulunan dinozora verilen isimdir. Kuzey Amerika ve Moğolistan’da, Protoceratopsidae familyasında yer alan buna benzer çeşitli dinozorlar bulunmuştur. Romer’in de belirttiği gibi, Protoceratops’a, hiç boynuzu olmadığı için, yanlış isim verilmiştir. Burun bölgesi boynuzumsu yapıdadır ve bazı bireylerin üzerinde buruşukluklar vardır. Evrimciler, bu tür canlılarda boynuzların gelişebileceğini düşünmektedirler; fakat bunu gösteren hiçbir geçiş formu bulunamamıştır. Üstelik, yukarıda da söylendiği gibi, Protoceratops, tüm boynuzlu dinozorların bulunduğu dönem olan Üst Kretase’de bulunmuşlardır. Gerçekten, evrimcilere göre, protoceratops dinozorlarının en ilkellerinden birisi olarak tanımlanan Leptoceratops gracilis, Kuzey Amerika’da son olarak bulunan dinozorlardan biriydi.49 Eğer Protoceratops boynuzlu dinozorların atası olsaydı, Üst Kretase’den daha yaşlı olduğu kabul edilen Alt ve Orta Kretase gibi jeolojik oluşumlarda bulunması gerekirdi. Sonuç olarak Protoceratops, boynuzlu dinozorların atası olduğu iddiası böylece boşa çıkmış olmaktadır.
Triceratops’un olası evrimsel çeşitleri konusunda, iki dinozorun sözü edilmektedir. Bunlar, artık tam gelişmemiş bir Triceratops50 olduğuna inanılan Sterrholophus ile patolojik bir Triceratops biçimi51 olan Diceratops’tur. Weishampel ve diğerler şöyle der:
Protoceratopsidler ve gerçek boynuzlu dinozorlar olan ceretopsidler arasında büyüklük ve göreli büyüme (allometric) özellikleri olarak keskin bir devamsızlık vardır; ve familyaların üyeleri arasında bir karışıklık yoktur.52
Toplam uzunluğunun yaklaşık üçte birine eşit kafa ve kalkana sahip olan Triceratops dinozorundan farklı olarak, Stegosaurus, boynuzsuz, küçük bir kafaya sahipti. Ancak Stegosaurus, bazı alışılmadık yapılara sahipti. Bu yapılar, kuyruk üzerindeki, her biri yaklaşık bir metre uzunluğunda olan çivi benzeri boynuzlar ile, boynu, gövdeyi ve kuyruğu boydan boya kaplayan plâkalardan oluşuyordu. Altı metre uzunluğunda dört ayaklı olan bu dinozorlar çivi benzeri yapılarını elbette ki kendilerini korumak için kullanıyorlardı; fakat plâkaların ne işe yaradığı, şüpheli bir konudur. Bazı insanlar, iskelete bağlarla bağlanmış halde bulunan plâkaların zırh görevi gördüğünü ileri sürmüşlerdir. Ancak, birbiri ardına gelen çift sıra düzeni ve plâkaların konum şekli, bu plâkaların, ısı değişimini sağlama görevleri olduğu görüşünü desteklemektedir. Ancak, plâka ve çivi yapılarının kademeli evrimsel kökenini gösteren bir geçiş formu dizisi bulunamamıştır. Bol miktarda bulunan Stegosaurus fosilleri, evrimden beklenenin tam tersine, yaratılış ile uyum göstererek, tümüyle gelişmiş biçimde ortaya çıkmaktadır.
Bir diğer alışılmadık dinozor çeşidi de, hadrosaur ya da ördek gagalı dinozordur. Bu iki bacaklı canlıların çok sayıda fosili Üst Kre